

서 론
블루베리는 안토시아닌과 폴리페놀 화합물을 다량 함유하여 항산화 활성이 높고 시력 보호 효과가 크다(Castrejón 등, 2008; Kalt 등, 2001; Prior 등, 1998). 이로 인해 블루베리의 재배 면적과 소비가 세계적으로 증가하고 있다. 우리나라에서 재배되는 블루베리는 주로 남부하이부쉬와 북부하이부쉬 품종인데, 남부하이부쉬는 저온 요구도가 낮아 남부 지방에서 노지 및 시설 재배가 늘고 있다. 이 품종은 환경 적응성이 높아 국내외 따뜻한 지역에서도 인기를 끌고 있다(Bañados, 2009; Finn 등, 2014). 특히 남부하이부쉬는 저온 요구도가 낮아 조기 수확을 위한 시설 재배에 유리하다(Baptista 등, 2006; Ciordia 등, 2002; Ogden과 van Iersel, 2009). 우리나라의 블루베리 시설 재배는 비가림, 무가온, 가온재배로 나누는데 남부지방에서 남부하이부쉬 품종은 주로 가온 재배로 이루어지며, 이는 이 품종이 저온 요구도가 낮고 고온 내성이 강하기 때문이다(Kim과 Yun, 2015; Moon 등, 1987). 가온재배에서는 7.2℃ 이하에서 200시간 이상의 저온 요구도를 충족한 후 가온을 시작한다(RDA, 2020). 일부 남부하이부쉬 품종은 연중 녹색을 유지하는 상록수 특성을 가지고 있다(Lyrene, 1997, 2008). 이러한 상록형 품종은 저온 요구도가 낮아 겨울이 따뜻한 지역에서 고생산성 재배가 가능하다(Bañados 등, 2009; Brazelton 등, 2013; Maatala 등, 2020). 상록형 재배는 기온을 높게 유지하고 양분을 지속적으로 공급해야 하지만, 국내 노지나 무가온 시설에서는 어렵다. 그러나 가온 시설에서는 저온 요구도가 낮은 ‘신틸라’ 품종을 이용한 상록 재배가 가능할 것으로 예상된다. 본 연구는 ‘신틸라’ 품종을 대상으로 가온 시설 내에서 상록 재배 가능성을 확인하기 위해 수행되었다. 상록형과 낙엽형 재배 방식을 비교 분석하여, 가온 전 온도와 양분 공급이 잎의 상태와 생장에 미치는 영향을 조사하였다.
재료 및 방법
1. 재배형태별 관리
본 연구는 2022년 10월부터 2023년 5월까지 경상남도 진주에 위치한 비닐온실과 노지에서 4년생 ‘신틸라’ 블루베리 나무를 대상으로 상록형과 낙엽형 재배 시험을 수행하였다. 각 재배 형태별로 5주씩 총 10주를 사용하였다. 나무는 모두 32L 원형 플라스틱 용기에 피트모스(NAROPEAT, EU)와 펄라이트(SJ Corp. China)를 8:2 비율로 혼합한 상토에 심겨졌으며, 열 간격 1.5m, 주 간격 1.0m로 배치되었다.
상록형 재배는 비닐온실 내에서 전 생육기간을 유지시켰으며, 일반적인 잎의 노화기인 10월부터 11월까지 무가온 상태로 개폐장치를 활용하여 낮 28℃, 밤 10℃ 이상으로 온도를 관리하고 12월 1일부터는 가온을 시작하였다. 일반적으로 낙엽형 재배는 수확이 끝난 후 비가림 형태의 온실에서 노지와 비슷한 환경으로 가온 전까지 보내지만 본 시험에서는 노지에서 11월 28일까지 관리하다가 11월 28일에 비닐온실로 옮겼다. 10월부터 11월 까지의 낮 평균온도는 21.8℃였지만, 야간의 최저온도는 10월 상순 12.1℃, 10월 하순 2.2℃, 11월 하순에는 –1.4℃까지 낮아졌다. 11월 28일에 낙엽형 나무는 비닐온실로 옮겨, 12월 1일부터 상록형과 동일한 온도 및 양분조건으로 재배하였다.
가온은 두 재배형태 모두 12월 1일에 시작하였는데, 이때 상록형 시험수는 개화 전 60일, 낙엽형은 개화 전 72일이었다. 겨울 동안 주야간 온도가 10℃ 이상 유지되도록 가온을 하고, 봄에는 추운 야간에만 가온하여 온도가 8℃ 이하가 되지 않도록 하였다. 양분은 Cheon 등(2018)이 제시한 블루베리 무가온재배용 표준양액(NO3-N 4.6, NH4-N 3.4, PO4-P 3.3, K 3, Ca 4.6, Mg 2.2mmol·L-1)을 양액기로 공급하였는데, 재배형태별로 공급시기를 달리하였다. 상록형은 2022년 수확이 끝난 후 계속 공급하였고, 낙엽형은 2022년 6월 하순(수확 종료 50일)까지 공급하고 이후 중단하였다가 가온 시작일인 12월 1일부터 공급하였다. 양액은 두 재배형태 모두 EC를 1.0으로 맞추어 일주일에 7회, 4L씩 공급하였다. 양액과 별도로 관수를 하였으며, 겨울에는 주당 하루 평균 2L로 하다가 봄이 되면서 점차 늘려 6월에는 4L 정도로 나무의 증산량을 고려하여 조절하였다. 두 재배형태 모두 과다착과를 방지하기 위하여 개화 전까지 결과지당 화총이 5개 이상이 되지 않도록 화총을 솎았고, 개화기에 벌을 방사하여 수분을 촉진하였다. 생장이 늦었던 2차 결과지의 화총은 솎거나 적과는 하지 않았다. 그 외 관리방법은 농촌진흥청 블루베리 재배법(RDA, 2020)에 준하였다.
2. 생육 조사 및 통계분석
엽록소함량은 2022년 10월 11일부터 12월 20일까지 SPAD-502(Konika Minolta, Japan)로 측정하였다. 신초경, 신초장, 주당 신초수는 2023년 2월 15일과 5월 15일에 각각 주당 10개의 신초를 대상으로 조사하였다. 3월 13일에는 나무별로 정상 결과지보다 40일 이상 늦게 발생하여 개화하는 2차 결과지수를 조사하였다.
수량은 3-5월 수확기에 5회로 나누어 수확한 후 주당 총 수량을 구하였다. 4월 10일에 수확한 과실 중에서 주당 50개씩 무작위로 골라 평균 과중을 조사하고 과실 적도 면에서 횡경을 측정한 후 모든 과실을 으깨어 착즙하고 굴절당도계(PR-100, Atago, Japan)로 가용성 고형물 함량을 측정하였다. 조사성적은 1주를 1반복, 처리별로 5반복으로 정리하여 R 통계프로그램(i386 4.1.0)을 이용하여 T-test로 두 재배형태를 비교 분석하였다.
결과 및 고찰
1. 잎의 변화
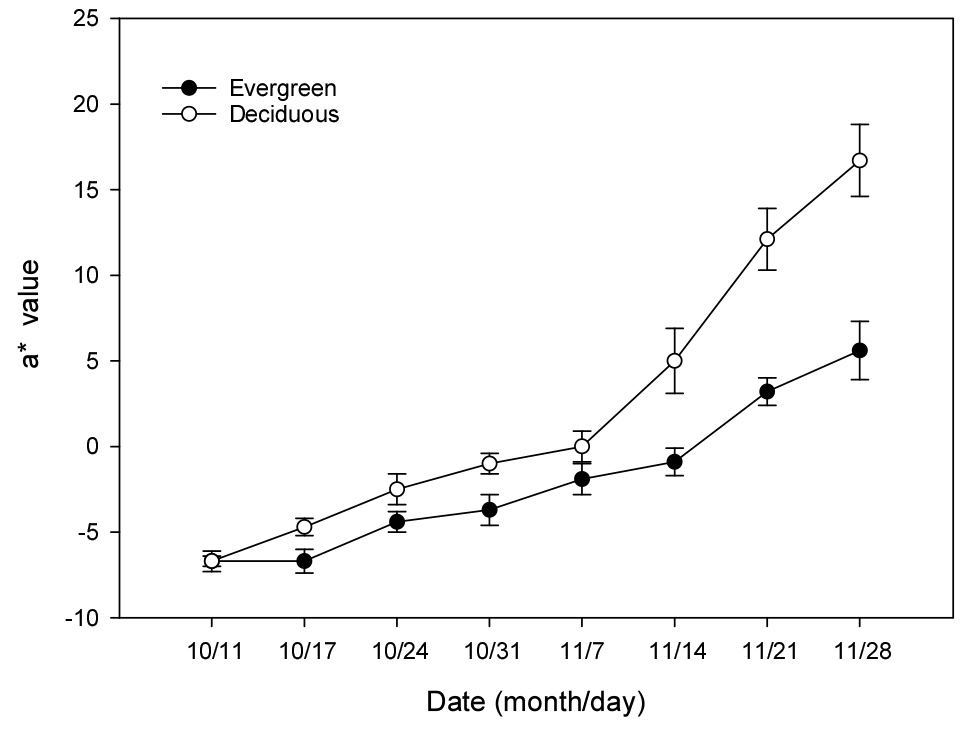
남부하이부쉬 블루베리 ‘신틸라’ 나무들을 상록형과 낙엽형으로 나누어 가을철 온도와 양분공급을 달리했을 때 잎의 녹색과 붉은색을 나타내는 Hunter a 값은 시간이 지날수록 재배형태에 따라 점진적인 차이를 보였다. a* 값은 온실 안에 있었던 상록형 나무 잎에서 10월 11일 –6.7(녹색)에서 11월 28일 5.6으로 붉은색이 약간 증가한 반면, 노지의 낙엽형에서는 –6.7에서 16.7로 붉은색 값이 상록형보다 2.9배로 높아져 단풍이 심하게 들었다(Fig. 1). 가을철 잎의 단풍은 노화와 함께 기온이 낮아지면서 엽록소가 감소하고 원래 있던 안토시아닌이나 카르티노이드 색소가 드러나기 때문으로(Mayta 등, 2019), 질소 양분의 공급이 적으면 잎의 노화는 더 빨리 나타난다(Choi 등, 2013)고 알려져 있다.
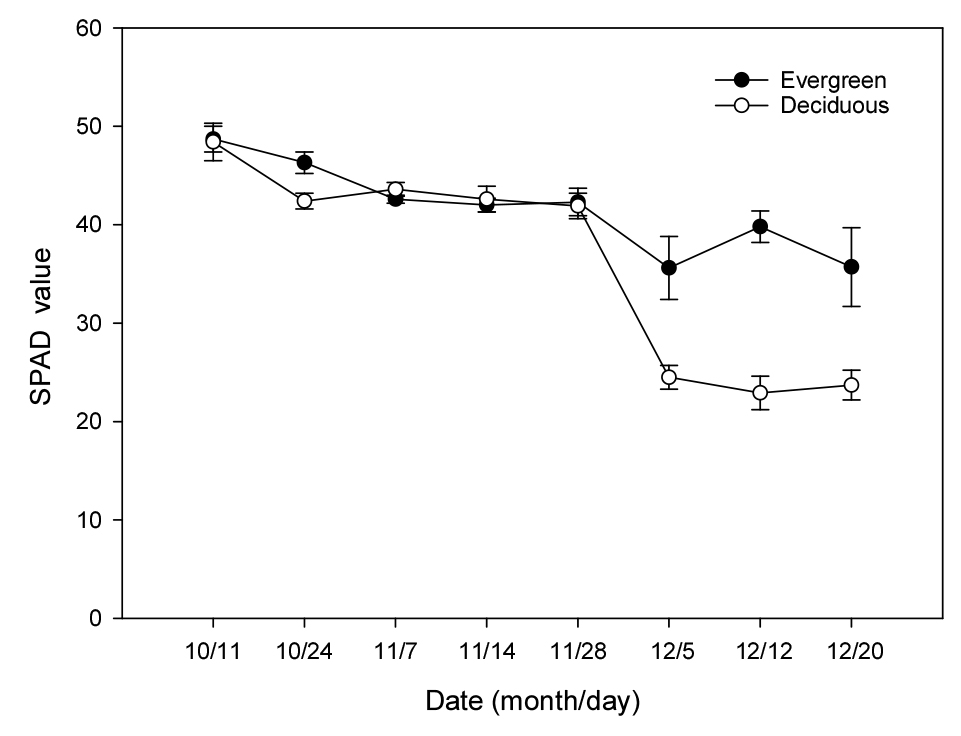
SPAD 값은 10월 11일에 상록형과 낙엽형이 각각 48.7, 48.4에서 기온이 낮아진 11월 28일에는 42 정도로 서서히 감소하여 a* 값의 변화(Fig. 1)와 달랐다(Fig. 2). 그러나 가온 시작 5일째인 12월 5일에는 상록형이 35.6인데 반해, 낙엽형은 24.5로 현저히 낮아져 잎의 광합성 기능이 급격히 감소하였음을 나타냈다. 12월 상순 이후 상록형의 SPAD 값이 낙엽형보다 높은 것은 잎이 건전하였고, 가온 전 양분공급으로 잎의 질소 성분이 많았기 때문(Choi 등, 2011)으로 이해된다. 가온과 양분공급이 시작된 이후 상록형의 엽록소 값이 점진적으로 증가하는 경향을 보였다. Fig. 3에 더욱 뚜렷하게 확인할 수 있었는데, 11월 28일에 단풍이 든 잎이 2월 10일에는 녹색으로 상당히 회복되었다. 이러한 결과로 볼 때 잎의 노화 과정에서 엽록소가 파괴었다가 다시 생성되는 지, 파괴되지 않고 존재하고 있다가 환경이 좋아지면 활성화되는 것인지에 대해서는 추가적인 조사가 필요할 것으로 여겨진다.

2. 수체 생장
진주지역에서 낙엽형과 상록형 재배 이듬해의 생육 상태를 조사한 결과는 Table 1과 같다. 신초직경, 평균신초장, 주당 신초수 등 신초생장은 과실비대 초기인 2월 15일과 수확인 끝난 5월 15일 모두 상록형이 낙엽형보다 왕성하여, 5월 15일 신초장은 상록형이 낙엽형의 10.1cm보다 52%나 길었고, 주당 신초수는 낙엽형의 187개보다 35% 많았다. 이는 전년도 11월까지 외기 온도 조건에서 재배한 낙엽형은 8월 이후 양분공급이 중단된 반면, 상록형은 지속적으로 양분이 공급되어 수체 내 저장양분 축적에 유리하였고, 배지 상토에도 비료 성분이 높게 유지되어 이듬해 양분흡수량도 많았을 것이다. 특히, 블루베리의 질소 영양이 많아지면 신초 생장은 현저하게 증가한다(Ballinger 등, 1963; Ballinger와 Kushman, 1966). 또한 낙엽형에 비해 상록형이 전년 10-11월 시설 내 기온이 높아 12월 이후 수체 생장이 빨랐던 탓도 있을 것이다.
1차 결과지는 작년 결과지에 화아분화가 되어 이듬해 과실이 맺히는 결과습성을 가지고, 2차 결과지는 당해연도 발생한 신초에서 꽃이 피고 과실이 맺히는 가지를 말하는데, 2차 결과지의 꽃수는 낙엽형에 비해 상록형이 6.8% 많았다. 신틸라의 2차 결과지는 당년 발생한 새가지 잎이 단일의 광주기를 인식하여(Lyrene, 1992), 꽃눈분화가 되어 개화한 것으로 수체의 양분상태와도 관련이 있다(Darnel 등, 1991; Reeder 등, 1998). 상록형 재배에서는 수체의 양분상태가 좋아 새가지의 꽃눈분화로 인한 2차 결과지 발생이 많았던 것으로 생각된다. 2차 결과지의 과실은 생장과 수확시기가 늦지만 과실품질이 정상과실과 차이가 없으므로(Cheon 등, 2023) 수량 확보에 유리하다.
연중 온실에서 잎을 녹색으로 유지했던 상록형은 노지에서 잎의 노화를 겪은 낙엽형보다 개화와 수확이 빨랐다. 상록형은 개화가 2월 3일에 시작되어 낙엽형보다 8일이 빨랐으며, 수확은 3월 23일 시작하여 11일 빨랐다(Table 1). 이는 신초생장에서 언급했듯이 낙엽형에 비해 상록형이 전년 10-11월 시설 내 기온이 높아 12월 이후 수체 생장이 빨랐던 탓이었을 것이다. 한편 대부분의 남부하이부쉬 품종들은 저온요구도가 200시간 이상 되어야 정상적인 개화가 된다(RDA, 2020). 그러나 본 시험에서 신틸라의 상록형 재배는 개화 전까지 10℃ 이상으로 관리되어 이러한 저온요구도가 충족되지 않았는데도 정상적인 개화가 이루어졌으므로, 이 품종의 저온요구도는 200시간보다 훨씬 낮은 것으로 판단되었다.
3. 과실 특성 및 수량
Table 2는 수확한 과실의 특성과 수량을 조사한 결과이다. 상록형의 과중은 2.4g, 횡경은 16.1mm로 낙엽형보다 각각 17%, 6%가 컸다. 상록형의 이러한 과실 크기 증가는 Table 1에 나타났듯이 증가한 영양생장에 따른 엽면적 확대로 동화산물의 과실 공급량이 많았기 때문(Maust 등, 1997)으로 생각된다. 그러나 가용성 고형물은 재배형태에 따른 유의적인 차이가 없어 상록형의 영양생장 증가(Table 1)나 조기 성숙이 과실 내 가용성 고형물 함량 증가로 반드시 이어지지 않았음을 알 수 있었다. 상록형의 주당 수량은 2.2kg으로 낙엽형보다 29% 많았는데, 과중 증가가 낙엽형 대비 17%인 점을 감안하면 과중보다 착과수가 수량에 영향을 준 것으로 여겨진다.
Table 1.
Responses of southern highbush blueberry ‘Scintilla’ in 2023 to the evergreen and deciduous growing types controlled with different temperature and nutrient during late autumn in 2022.
| Growing type |
Shoot diameter (mm) |
Shoot length (cm) |
No. of shoots per bush |
Flowers on secondary bearing shootz (No./bush) | Flowering period | Harvesting period | |||
| Feb. 15 | May 15 | Feb. 15 | May 15 | Feb. 15 | May 15 | ||||
| Evergreeny | 1.5 | 2.1 | 11.2 | 20.9 | 122 | 285 | 266 | Feb. 3 to Feb. 11 | Mar. 23 to May 10 |
| Deciduousx | 1.4 | 1.7 | 4.8 | 10.1 | 78 | 187 | 248 | Feb. 11 to Feb. 20 | Apr. 3 to May 15 |
| Significance | NS | * | ** | * | ** | * | * | ||
Table 2.
Berry characteristic of southern highbush blueberry ‘Scintilla’ in 2023 as affected by the evergreen and deciduous growing types controlled with temperature and nutrient supply during late autumn in 2022.
| Growing type |
Weight (g) |
Diameter (mm) |
Soluble solids (°Brix) |
Yield (kg/bush) |
| Evergreenz | 2.4 | 16.1 | 12.8 | 2.2 |
| Deciduousy | 2.0 | 15.1 | 12.5 | 1.7 |
| Significance | * | NS | NS | * |
