서 론
국내에서 생산되는 토마토(Solanum lycopersicum)의 대부분은 온실에서 재배되고 있으며(RDA, 2023), 특히 다른 과채류에 비해 재배가 용이한 토마토는 수경재배 방식이 확산됨에 따라 전국적으로 재배되고 있다(RDA, 2020). 스마트 온실의 도입은 건강식으로 주목받는 토마토의 꾸준한 수요에 대응할 수 있는 안정적인 공급망 역할을 하고 있을 뿐만 아니라 노동력 절감을 통해 생산비 절약에도 도움이 된다. 또한, 스마트 온실은 작물이 자랄 수 있는 최적의 환경을 조성하여 일정한 품질 관리와 생산성 향상에도 효과적이다(Shamshiri 등, 2018). 최근에는 토마토 이외에도 딸기, 오이, 파프리카 등 여러가지 과채류들도 스마트 온실에서 재배하는 방식으로 변화하고 있다.
토마토의 품질은 크기, 무게, 경도, 색상과 같은 외형적인 요소를 통해 판단된다. 이러한 요소들은 기온, 일사량 등 여러 가지 환경 요인에 의해 영향을 받는다. 그 중 색상은 성숙 단계를 구분하고 수확 시기를 결정하는 가장 직관적인 요인이다. 과실의 색상을 활용한 성숙 단계 구분은 개인의 주관에 따라 달리 분류될 수 있지만 비파괴적이고 간단한 방식이기 때문에 대부분의 농가에서 수확 시기를 결정하기 위해 활용되는 방법이다. 착과 초기에 토마토는 초록색이며 재배되는 동안 표면에 빨간색의 비율이 늘어나기 시작하고 완숙기에는 완전한 빨간색으로 변한다. USDA에서는 토마토 표면의 빨간색 비율을 기준으로 성숙도를 6단계로 구분하는 것을 제안하였다(USDA, 1991). 이 기준에 따라 분류된 토마토는 초록색에서 빨간색으로의 변화를 나타내는 색상 값에서 뚜렷한 차이를 보였다(López Camelo와 Gómez, 2004).
토마토는 생육 과정에서 나타나는 과실의 색상 변화가 매우 뚜렷하여 이를 활용한 많은 연구들이 수행되었다. 실제로 토마토의 색상은 성숙도와 경도, 영양 상태를 평가하는 요소로 활용되고 있다(Hertog 등, 2004; Schouten 등, 2007). Tijskens와 Evelo(1994)는 수확 후 토마토의 색상 변화를 로지스틱 모델을 활용하여 보관 온도가 토마토 성숙도에 미치는 영향을 분석하였다. 비슷한 방법으로 일본에서 재배되는 세 가지 토마토 품종을 활용하여 수확 후 보관 온도에 따른 색상 변화를 모델링하고 효과적인 품질 관리와 공급 과정에서의 손실을 최소화할 수 있는 관리 방안을 제시하였다(Ciptaningtyas 등, 2022). 이 외에도 색상을 이용하여 성숙도를 나타내는 수치적 기준에 대한 연구도 진행되었다. Sharma 등(2020)은 토마토의 성숙도를 Ripening index (RI%)와 Red tomato(RT%)를 이용하여 수치적으로 계산하고 두 기준을 비교하였다. 두 기준 모두 토마토의 색상을 활용하여 계산되었고 RI가 토마토의 성숙도 변화를 나타내는데 RT보다 민감하게 반응한다는 결과를 보였다. 최근에는 딥러닝 기술과 이미지를 활용하여 토마토의 개체 인식과 성숙도 판단을 통해 수확 시기와 수확량을 예측하는 모니터링 로봇 연구도 진행되고 있다(Seo 등, 2021). 딥러닝 기술을 활용한 연구에서도 색상 변화는 숙도를 판단하고 수확 시기를 예측하기 위해 중요한 변수로 활용되고 있다.
온실에서 재배되는 토마토를 효율적으로 관리하기 위해서는 생육 과정 전반에 걸친 분석을 통해 온실 내 환경적인 요인의 영향을 파악해야 한다. 기존의 연구들은 수확 후의 관리와 작업의 자동화를 위한 숙도 예측 및 분류 모델에 관한 연구에 집중되어 있다. 따라서, 본 연구에서는 토마토 열매가 성숙되는 동안의 색상 변화를 수학적 모델을 이용하여 구축하고, 이 모델의 결과를 활용하여 재배 시기에 따른 성숙 과정의 차이를 확인하고자 하였다. 이를 위해 토마토의 색상 변화를 로지스틱 성장식을 이용하여 모델링하고, 재배 시기에 따라 완숙 토마토가 되기까지 걸린 시간의 차이를 수치적으로 비교하였다. 또한, 재배 시기의 온실 내 환경 요인을 수집, 분석하여 과실이 성숙되는 과정에 영향을 미치는 요인을 확인하고자 하였다. 본 연구는 토마토 과실의 성숙 과정을 관찰하여 재배 시기에 따라 성숙 속도에 차이가 있는지 확인하고자 진행된 실증적 연구로써, 착과 후부터 수확기에 걸친 토마토의 성숙 과정을 설명하기 위한 모델링이 수행되었다. 이 연구 결과를 바탕으로 적절한 생육 환경을 유지하는 온실 재배에서도 재배 시기를 고려한 온실 환경 제어와 관리의 중요성을 강조하기 위한 연구 결과로 활용될 수 있을 것으로 생각되었다.
재료 및 방법
1. 실험 설계
본 연구는 국립농업과학원에 위치한 반밀폐형 첨단디지털온실(전라북도 완주군)에서 수행되었으며, 온실의 동고와 측고는 각각 8.3m, 7.0m, 면적은 960m2이었다. 온실 환경 제어는 온실 중앙부 작물의 생장점 부근에 설치된 백엽상 데이터를 기반으로 이루어졌다. 온실의 내부 온도는 15℃-25℃ 범위로 유지하였으며, 고온기의 경우 냉방부하에 따라 자연환기, 강제환기, 포그시스템, 히트펌프를 순차적으로 가동하여 냉방하였고, 저온기의 경우에는 히트펌프를 이용하여 난방하였다. 또한, 온실 내부의 온도 유지를 위해 온실 내부와 외부에 설치된 차광막을 이용하였다. 차광막의 경우 여름에는 일사량이 600-700W/m2이상이 관측되면 작동시켰으며, 겨울에는 작동시키지 않았다. CO2 농도는 공급기를 이용하여 400-940ppm 범위로 유지되도록 하였다.
본 연구에서는 상향식 유인 방식으로 재배된 데프니스 품종(Dafnis, Syngenta Korea, Seoul, Korea)의 토마토를 실험 재료로 활용하였으며, 토마토는 코이어 배지(100×20×10cm, Daeyoung GS, Daegu, Korea)에서 재배되었다. 야마자키 토마토 배양액을 사용하여 재배하였으며, EC는 2.5dS·m-1, pH는 5.8로 관리하였고, 여름과 겨울 모두 동일한 농도의 양액을 사용하였다. 재배 시기에 따라 온실 재배 토마토의 생육 과정에 차이가 있는지 비교하기 위해 같은 장소에서 다른 기간에 재배된 토마토를 관찰하였다. 겨울에 재배된 토마토는 2022년 02월 14일부터 2022년 03월 31일까지 46일 동안 관측하였고, 20개의 샘플을 선정하였다. 여름에 재배된 토마토는 2022년 8월 11일부터 2022년 10월 4일까지 55일 동안 24개의 샘플을 관측하였다. 실험에 사용된 토마토는 10-13번째 화방에서 무작위로 선정되었으며, 화방당 착과수는 3-5개이었다.
토마토의 색상 변화는 과실이 완숙기에 도달할 때까지 2-3일 간격으로 측정하였다. 토마토의 색상 변화를 측정하기 위해 사용된 과실의 초기 평균 둘레는 11±1cm이었고, 실험 종료일 측정된 토마토 과실의 평균 둘레는 22±1cm이었다. 일반적으로 토마토는 후숙 과정을 고려하여 표면에 붉은색이 나타나기 시작할 때 수확하지만, 본 연구에서는 온실 재배 토마토의 생육 과정을 연구하기 위해 완숙기에 도달할 때까지 수확하지 않고 색상 변화를 관찰하였다. 토마토의 표면 색상은 휴대용 색도계(CR20, KONICA MINOLTA, Tokyo, Japan)를 이용하여 CIELAB 색 표시계를 측정하였다. CIELAB 색 표시계는 세 가지 색 좌표 L*, a*, b*를 활용하여 사람이 색상을 인식하는 방식과 비슷한 방법으로 색 공간을 표현한다(Westland, 2003). L*은 명도를 나타내는 값으로 0에서 100 사이의 값을 가진다. L*값이 0이면 검은색을, 100은 하얀색을 나타낸다. 또한, a*와 b*는 –128에서 127 사이의 값을 가지며 각각 초록색에서 빨간색, 파란색에서 노란색을 나타낸다. a*값이 음수이면 초록색, 양수이면 빨간색에 가까우며, b*는 음수이면 파란색, 양수이면 노란색으로 나타난다. 토마토의 CIELAB는 각 샘플마다 세 번 반복 측정하였고 이들의 평균값을 분석에 활용하였다. 토마토의 색상은 꼭지와 배꼽을 기준으로 과실의 중심에 해당하는 위치를 3반복 측정하여 평균으로 나타내었으며, 측정 위치를 표시하여 항상 동일한 위치에서 CIELAB를 측정하였다.
본 연구에서는 재배 시기에 따라 온실 내부의 환경이 토마토의 생육 과정에 미치는 영향을 확인하고자 추가적으로 센서를 설치하여 토마토의 색상이 관측되는 같은 기간 동안 온실 내부의 환경 데이터를 수집하였다. 환경 데이터는 온도-습도-기압센서(Web-Thermo-Hygrobarometer, W&T, Wuppertal, Germany)와 일사량센서(SMP10-V, KIPP & ZONEN, Delft, Netherlands)를 이용하여 측정하였으며, Fig. 1과 같이 온도-습도-기압센서는 토마토 과실 위치에 설치하였고 일사량센서는 작물이 없는 복도에 설치하여 내부 일사량을 측정하였다.

2. 로지스틱 성장식
로지스틱 성장식을 활용한 모델은 인구증가, 암세포나 기생충 같은 생물의 증식, 식물이나 과실의 성장 및 수확량 예측을 위한 연구 등 다양한 분야에서 활용되고 있다(Benzekry 등, 2014; Fernandes 등, 2019; Lee 등, 2017; Tsoularis와 Wallance, 2002). 로지스틱 성장식은 무한히 증가하는 형태의 지수함수 성장모델과 달리 시간이 지남에 따라 일정한 값으로 수렴하는 시그모이드(sigmoid) 형태를 보인다. 이러한 형태는 제한된 자원과 환경적 요인으로 인해 무한히 성장하지 못하는 현실의 성장 과정을 반영하고 있다. 대부분 작물의 성장은 최대값에 도달하기 전 느린 속도로 증가하다 빠르게 증가하며, 그 이후에는 증가 속도가 느려져 결국에는 성장이 둔화하는 형태를 보인다. 따라서 이러한 성장 패턴을 설명하기 위해 로지스틱 성장식과 같은 시그모이드 형태의 성장곡선이 주로 활용되고 있다.
본 연구에서는 시그모이드 형태의 변화를 보인 토마토의 a*값을 로지스틱 성장식을 이용하여 모델링하였다. 토마토는 미숙과에서 완숙과로 진행되는 동안 초록색에서 빨간색으로 색상이 변화한다. 따라서, 이러한 색상 변화를 활용하여 토마토 과실이 익은 정도를 판단할 수 있고 이때의 a*값을 활용하여 토마토 과실이 성숙하는 과정을 나타내는 모델을 구축하고자 하였다. 모델은 아래 Eq. (1)식과 같이 4개의 모수를 가지는 로지스틱 성장식을 활용하였다.
위 식에서 는 모델의 설명변수로 측정일이 사용되었다. 표면 색상이 처음 관측된 날을 1일로 하여 색상이 관측된 날의 일수를 계산하여 사용하였다. 첫번째 모수 는 초기 a*값이 음수임을 고려하여 추가된 모수로 관측 초기에 일정하게 유지되는 a*값을 나타낸다. 두번째 모수 는 시간이 지남에 따라 모델이 수렴하는 값인 최대한계수용능력(carrying capacity)을 계산하는데 활용된다. 로지스틱 모델에서 최대한계수용능력은 시간이 지남에 따라 모델이 수렴해 가는 값으로 본 연구에서 최대한계수용능력은 완숙기에 도달한 토마토가 수렴하는 a*값으로 로 계산된다. 세번째 모수 는 모델의 변곡점(inflection point)을 나타낸다. 변곡점은 모델의 성장률이 최대인 점으로, 로지스틱 성장식에서는 이 점을 중심으로 함수가 대칭의 형태를 가지는 특징이 있다. 마지막 모수 는 성장률과 관련 있는 모수이다.
로지스틱 모델과 같은 수리적 모델을 활용한 성장모델은 모수 값을 활용하여 모델 간 비교와 정량적인 평가가 가능하다는 장점이 있다. 또한 이 모델은 성장률의 변화를 나타낼 수 있는 5개의 임계점(critical points)을 추론하여 모델값의 변화를 설명할 수 있다. 5개의 임계점은 절대가속점(Absolute acceleration point, AAP), 최대가속점(maximum acceleration point, MAP), 변곡점(inflection point, IP), 최대감속점(maximum deceleration point, MDP), 점근감속점(asymptotic deceleration point, ADP)이다. 먼저 절대가속점 이전에는 증가율의 가속이 매우 느려 모델값이 거의 증가하지 않는다. 따라서 실제 모델값의 증가는 이 점 이후로 시작된다. 다음으로 최대가속점은 모델값의 증가율이 최대가 되는 점을 나타낸다. 앞서 언급했던 변곡점은 증가율이 최대이며 증가율의 가속이 0인 점으로, 이 점 이후에는 모델값의 증가가 둔화한다. 최대감속점은 모델 증가율의 가속이 최소가 되는 점을 나타낸다. 점근감속점은 성장률이 0으로 모델값이 더 이상 변화하지 않는 점을 의미한다. 이러한 임계값은 로지스틱 성장식의 미분함수 값을 0으로 만족하는 점에서 찾는 과정에서 추론되며 절대가속점과 점근감속점, 최대가속점과 최대감속점이 쌍을 이룬다. 절대가속점과 점근감속점은 3차 미분함수, 최대가속점과 최대감속점은 4차 미분함수를 통해 계산된다(Korkmaz 등, 2018). 이러한 로지스틱 모델의 특징을 활용하여 토마토의 성장에 걸리는 시간을 연구하고 수확기를 예측하였다(Fang 등, 2022; Silva 등, 2021).
본 연구에서는 재배 시기에 따라 모델의 모수에 차이가 있는지 분석하기 위해 Eq. (1)에 재배 기간을 구분하기 위한 더미변수 를 추가하여 아래 Eq. (2)와 같이 모델을 수정하였다. 아래와 같은 모델은 재배 시기에 따라 각각의 모델을 수립하지 않고도 모델을 수립할 수 있고, 모수의 유의한 차이를 비교하기 위해 유용하게 활용된다.
자료 분석과 모델 구축을 위해 R 프로그램(R Core Team, Vienna, Austria)을 사용하였다. 토마토의 색상 변화 자료에 로지스틱 성장식을 적용하고 모수를 추정하기 위해 stats 패키지(version 4.3.1)의 nls를 활용하였다. nls는 비선형방정식 모델을 적용을 위해 활용된 함수이고, 로지스틱 성장식의 모수는 수치해석적 방법인 Gauss-Newton algorithm을 활용하여 추정되었다.
결과 및 고찰
1. 토마토의 표면 색상
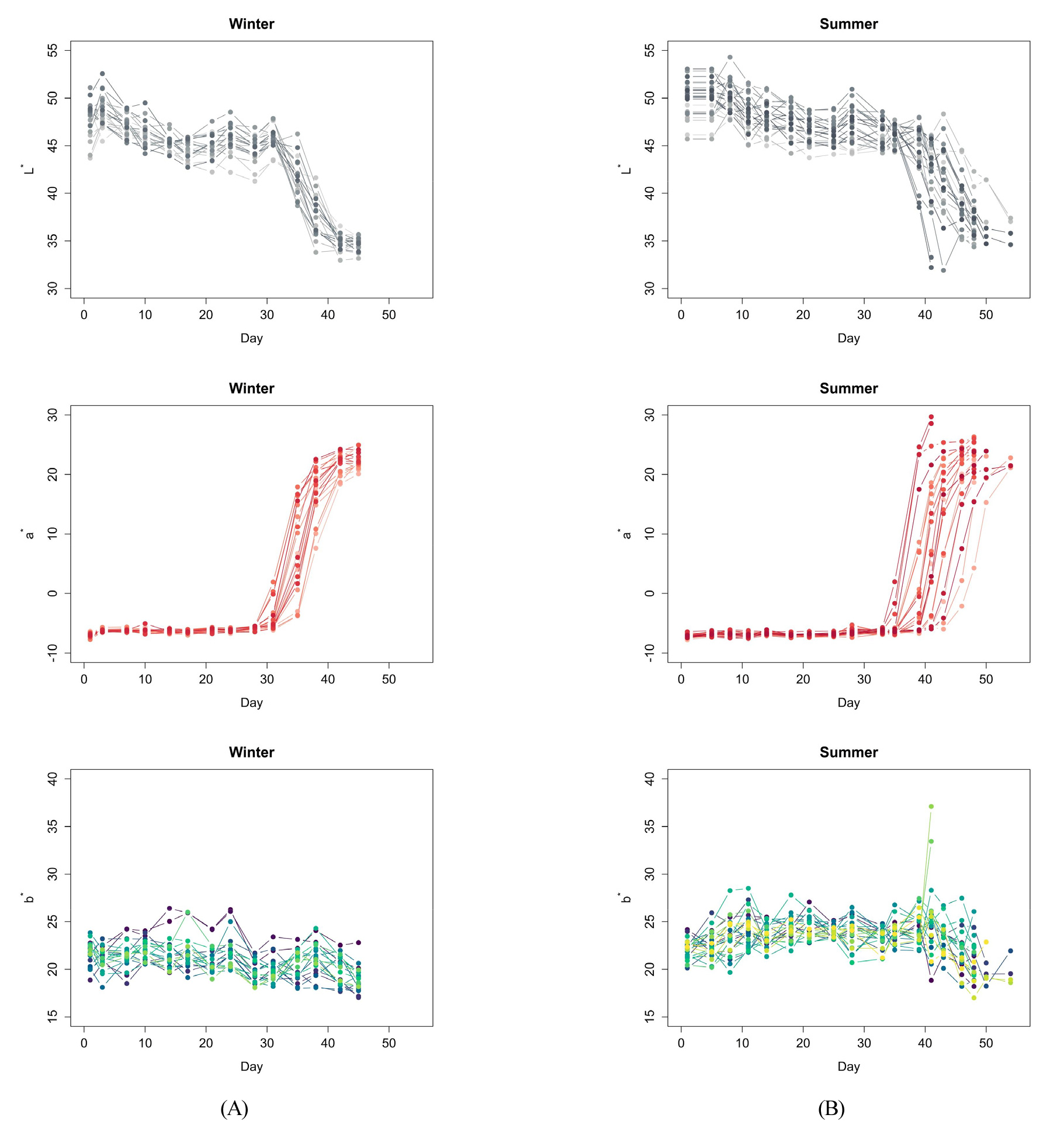
시간에 따른 토마토의 색상 L*, a*, b*를 관찰한 결과를 Fig. 2에 나타내었다. 토마토의 성숙 과정에서 측정한 세 가지 색상의 변화는 여름과 겨울에 재배된 토마토에서 비슷한 양상을 보였다. 밝기를 나타내는 L*값은 착과 초기에 45에서 55 사이의 값을 보이다 시간이 지남에 따라 감소하는 경향을 보인다. 이후 완숙기에 도달하면 35 근처의 값을 가진다. b*값은 측정 초기 20에서 30 사이의 값을 가지며 시간이 지나도 큰 변화를 보이지 않았다. 일부 여름에 재배된 토마토에서 30을 넘는 급격히 증가한 b*값이 관측되었는데, 이 경우의 토마토는 다른 토마토에 비해 빨리 완숙기에 도달한 토마토로 약 5일에서 10일 정도 빨리 수확된 샘플이다. 마지막으로 초록색에서 빨간색으로의 변화를 나타내는 a*값은 시그모이드 형태를 보였다. 측정 초기에 a*값은 –5에서 –10 사이의 일정한 값으로 유지되다 급격하게 증가하는 과정을 거쳐 20과 30 사이의 일정한 값으로 수렴하였다. 이러한 변화는 로지스틱 성장식을 활용하기 위한 변수로써 적절한 특징을 보이며, 토마토 과실의 성숙 과정을 나타내기 위한 모델을 구축하기 위해 활용될 수 있다.
재배 시기와 상관없이 비슷한 형태의 색상 변화가 나타났지만, 여름에 재배된 토마토에서는 겨울에 재배된 토마토보다 색상 변화의 변동성이 크게 나타났다. 이 현상은 특히, a*값에서 확연한 차이를 보인다. 겨울에 재배된 토마토에서는 각 샘플 마다 a*값이 증가하기 시작한 시기가 비슷하게 나타난다. 그러나, 여름에 재배된 토마토는 샘플 마다 a*값이 증가하는 시기에 차이가 있음을 알 수 있다. 토마토 과실이 성숙하는 과정에서 나타난 색상 변화는 토마토의 성숙도를 평가하기 위한 기존 연구 결과와 유사한 경향을 보였다(López Camelo와 Gómez, 2004; Tijskens와 Evelo, 1994). 특히, 시그모이드 형태의 a*값은 초기에는 빠르게 성장하고, 이후 완만한 성장 속도를 보이는 토마토의 성숙 과정을 효과적으로 나타내고 있다고 알려져 있다(Ciptaningtyas 등, 2022). 따라서 토마토의 색상 변화를 활용하여 성숙 과정을 모델링 하기 위해서 a*값을 활용하는 것이 효과적이라고 사료된다.
2. 로지스틱 모델 결과
본 연구에서는 로지스틱 성장식을 이용하여 겨울과 여름에 재배된 토마토의 a*값의 변화를 나타내는 모델을 구축하였다. 이 모델은 토마토 과실이 성숙하는 과정을 나타내고, 모델의 모수 값을 활용하여 재배 시기에 따라 토마토 과실이 완숙기에 도달하는 과정에 걸린 시간의 차이를 비교한다. 아래 식 Eq. (3), (4)는 겨울과 여름에 재배된 토마토의 시간에 따른 a*값의 변화를 나타내는 식이다. 모델식으로 추정된 a*값의 변화와 관측된 a*값을 Fig. 3에 나타내었다. Fig. 3에서 하늘색은 겨울, 분홍색은 여름에 재배된 토마토를 의미하며, 점선은 로지스틱 성장식으로 추정한 a*값이고 점은 관측된 a*값이다. 검정색 점선은 각 모델에서 변곡점을 나타내고 있다.
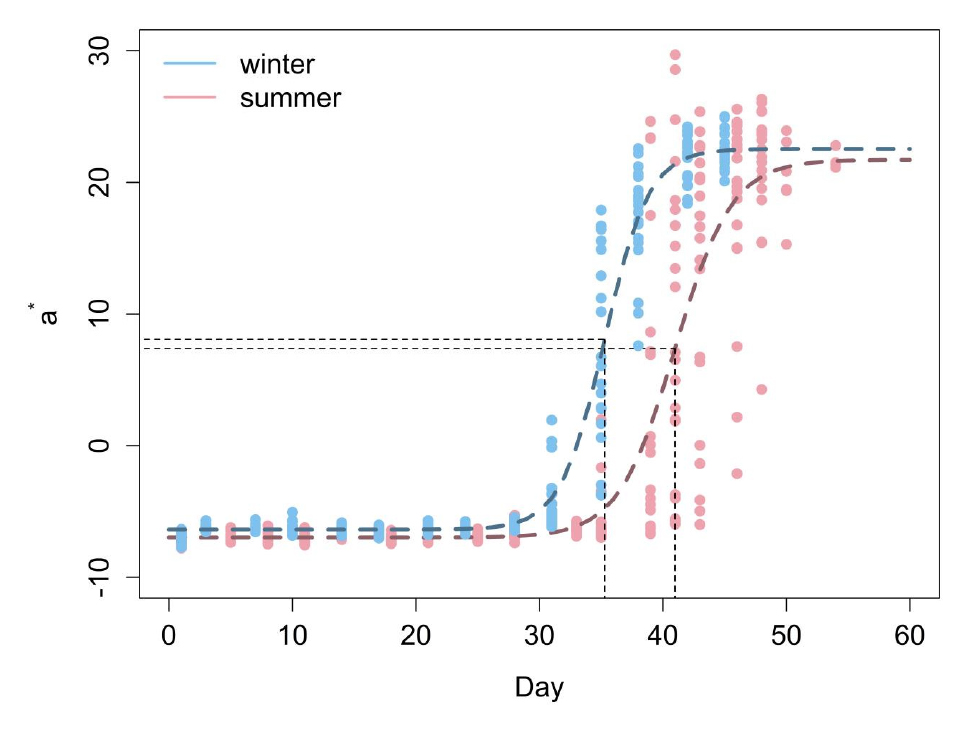
로지스틱 성장식으로부터 계산된 a*값의 초기값은 겨울에는 –6.3774, 여름 –6.9748로 겨울과 여름에 차이를 보이지 않았다. 이는 관측으로 얻어진 초기 토마토 과실의 a*값과 비슷한 값을 나타낸다. 한계수용능력값은 겨울에는 22.5480, 여름에는 21.7207로 완숙기에 도달했을 때 토마토의 a*값은 역시 재배 시기와 상관없이 같은 값으로 수렴함을 알 수 있었다. 이 값 역시 관측된 토마토에서 수렴하는 a*값과 비슷한 값을 보인다. 마지막으로 변곡점은 겨울에는 35일, 여름에는 40일로 계산되어 a*값의 변화율이 최대가 되는 시기는 재배 시기에 따라 차이가 있었다.
로지스틱 성장식에서는 모델값의 변화가 나타나는 시점이 변곡점을 중심으로 대칭으로 나타나는 특징을 가진다. 즉, 위와 같이 모델의 초기값과 최대한계수용능력 값이 같은 두 로지스틱 모델의 경우, 모델값이 변화하는 과정에 걸리는 시간을 비교해 볼 수 있다. 따라서 이 시간은 로지스틱 성장식의 임계값을 통해 추정해 볼 수 있다. 본 연구에서 활용한 로지스틱 성장식의 5개 임계점의 값을 계산한 결과를 Table 1에 나타내었다. a*값이 변화하는 데 걸리는 시간은 토마토 과실이 미숙과에서 완숙과가 될 때까지 걸리는 시간으로 설명될 수 있다. 모델값이 변화하는데 시간은 로지스틱 모델식에서 최대가속점과 최대감속점과 차이로 계산되고. 그 결과, 여름에 재배된 토마토의 a*값의 색상 변화는 약 10일, 겨울에 재배된 토마토는 약 8일에 동안 나타나고 있음을 알 수 있다. 본 실험에서 관측한 토마토의 경우, 과실이 성숙하는 데 걸린 시간이 여름보다 겨울에 짧게 나타났다. 이는 겨울에 재배된 작물이 여름에 재배된 작물보다 빨리 성숙 된다는 일반적이지 않은 결과를 나타내고 있다. 그러나, 이 결과는 토마토의 색상 측정을 위해 진행한 실험이 여름보다 겨울에 더 짧은 것과 비슷한 결과를 보인다. 토마토가 완숙기에 도달할 때까지 실험을 진행한 기간은 여름에는 55일, 겨울에는 46일이 걸렸다.
로지스틱 모델은 동물이나 식물의 성장 과정을 나타내기 위해 일반적으로 활용되는 비선형 성장식이다(Fernandes 등, 2019; Ribeiro 등, 2018). 이 모델은 작물의 재배 과정에서 나타난 초장, 엽면적 지수, 줄기, 잎, 열매 등의 생육 지표의 변화를 모델링 하거나 수확량 예측을 위한 모델로 활용되고 있다(Fang 등, 2022; Sari 등, 2019). 본 연구에서는 이러한 로지스틱 성장식을 토마토의 색상 변화를 모델링하는데 활용하고, 모델의 함수적 특징과 임계값을 활용하여 과실이 성숙하는 과정에서 걸린 시간의 차이를 비교하였다. 로지스틱 성장식은 모수를 활용하여 생물학적 해석이 가능하다고 알려져 있다(Mischan과 Pinho, 2014). 따라서 성장식에서 추론되는 임계값은 모델값이 증가하고 감소하는 시기를 의미하며(Korkmaz 등, 2018), 이를 이용하여 토마토의 성숙 속도가 변화하는 시기를 추론하였고 겨울과 여름에 재배된 토마토에서 과실이 성숙하기까지 걸린 시간의 차이가 있음을 확인하였다.
3. 온실 환경 및 생육 데이터 분석
재배 시기가 다른 토마토의 생육 과정에 영향을 미치는 온실 내 환경 요인을 확인하고자 온도와 일사량의 차이를 비교하였다. 토마토가 재배되는 동안 온실 내 일 평균 온도와 적산온도를 Fig. 4에 나타내었다. 일 평균 기온과 적산 온도 모두 겨울과 여름에 따라 큰 차이를 보이지 않았으며, 이는 실험 대상 온실이 반밀폐형 온실이며 냉난방 시설이 잘 갖추어져 있어 온도가 적절하게 유지되었기 때문이라고 판단된다. 그러나, 온도 변화가 나타나는 범위는 겨울보다 여름에 크게 나타나고 있음을 알 수 있다. Table 2에서와 같이 겨울 토마토가 재배되는 동안 평균 기온은 20.83℃, 여름 토마토의 평균 기온은 21.43℃로 여름 토마토의 재배 기온이 0.6℃ 높게 나타났다. 토마토의 생육에 적합한 온도는 주간 21℃-29.5℃, 야간 15.5℃-21℃로 알려져 있다(RDA, 2020). 수집된 온도에 따르면 토마토가 재배 기간 동안 온실 내 온도는 생육에 적합한 온도로 유지되었음을 알 수 있다.
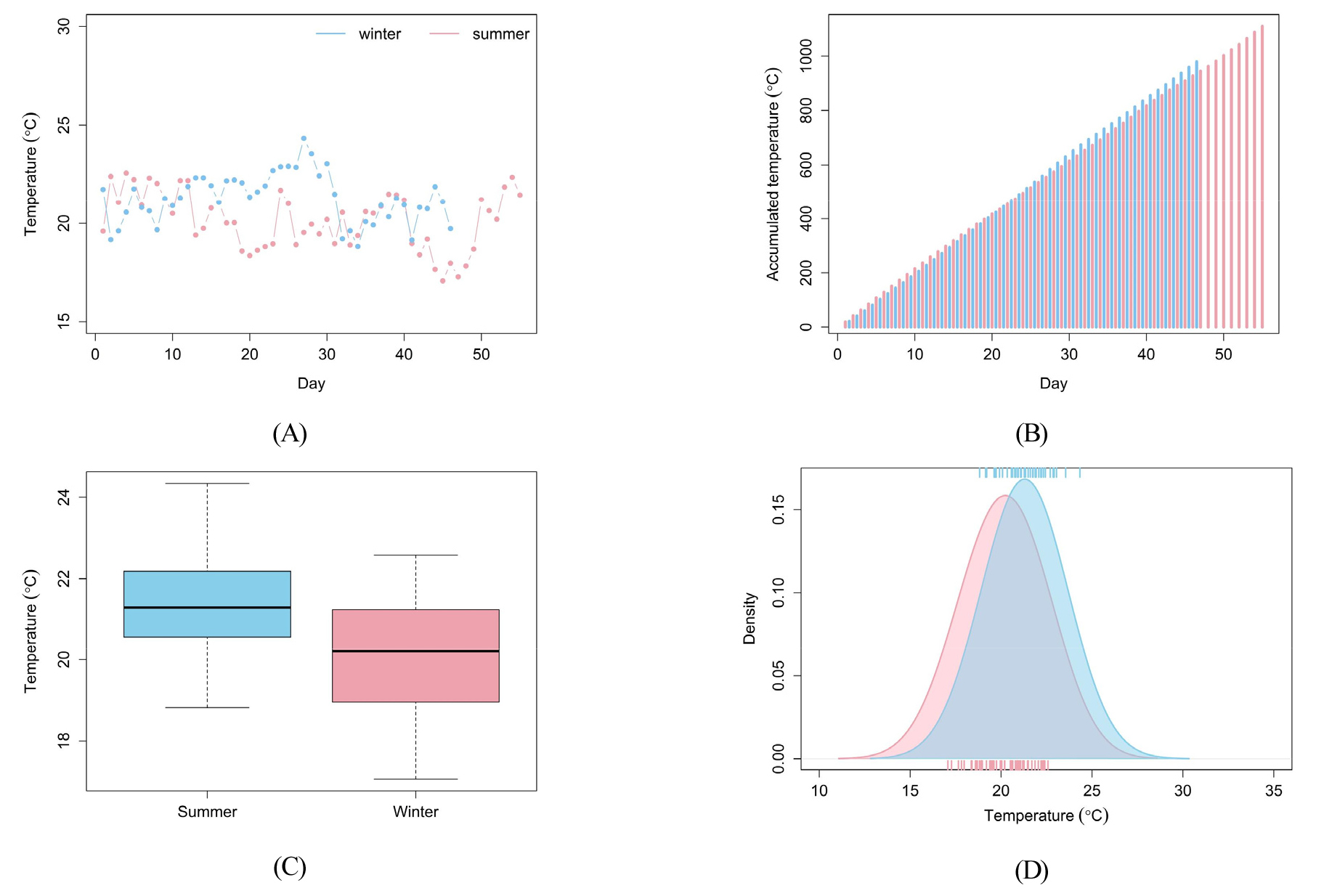
Table 2.
The summary statistics of temperature on winter (February 14, 2022 to March 31, 2022) and summer (August 11, 2022 to October 4, 2022).
|
Temperature (℃) | Growing season | |
| Winter | Summer | |
| Min | 18.82 | 17.07 |
| Q1 | 20.75 | 19.19 |
| Median | 21.43 | 20.64 |
| Mean | 20.83 | 21.43 |
| Q3 | 22.07 | 22.05 |
| Max | 24.33 | 26.00 |
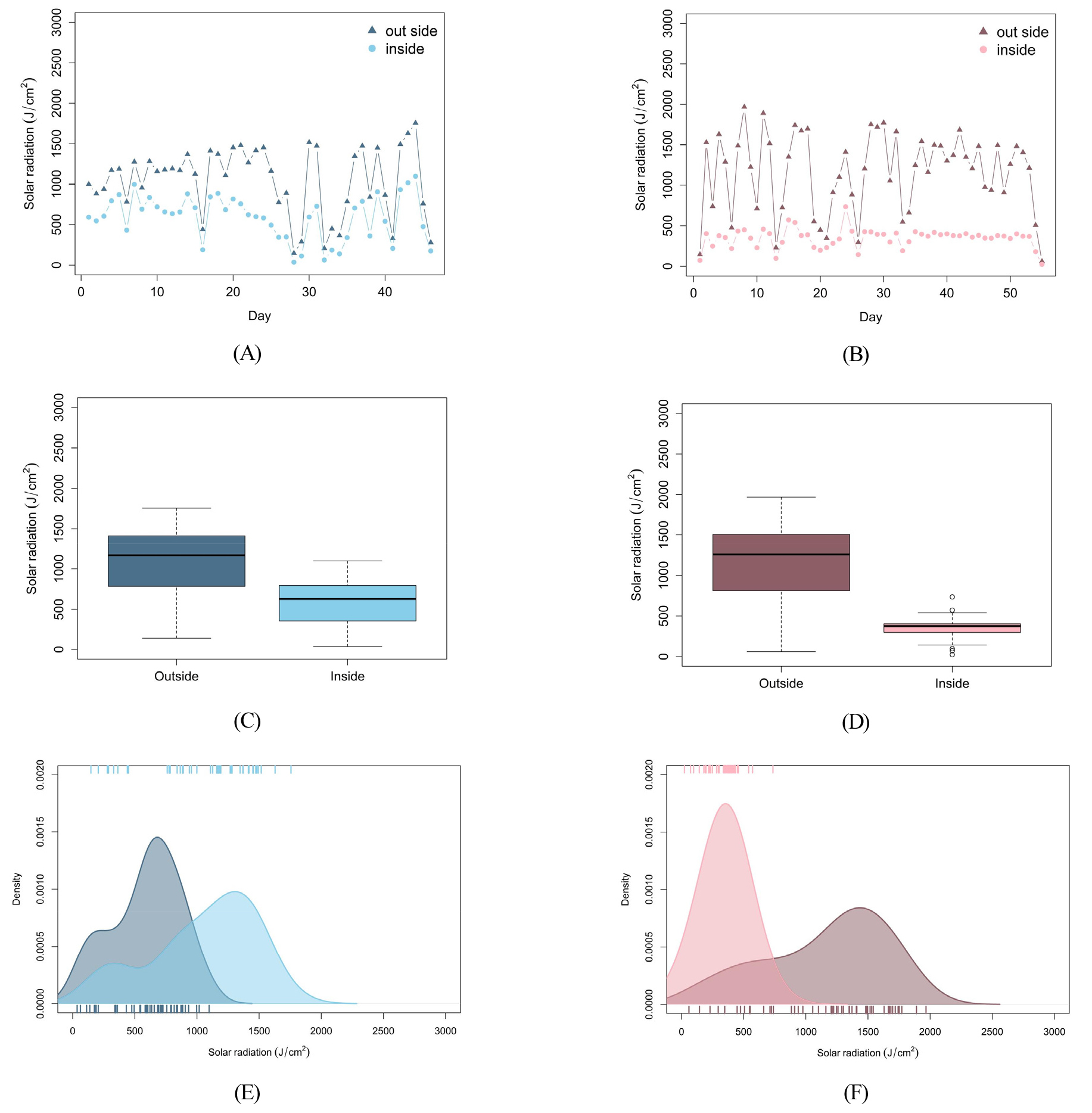
다음으로 토마토가 재배되는 기간에 측정된 외부와 내부 일사량을 Fig. 5에 나타내었다. 일사량은 일별 일조 시간에 해당하는 시간 동안 측정된 일사량을 누적하여 합한 적산일사량을 의미한다. 외부 일사량은 겨울보다 여름에 높았으나 내부 일사량은 여름보다는 겨울에 높게 나타났다. Table 3에서와 같이 외부 일사량은 여름에는 1161.8J/cm2, 겨울에 1051.1J/cm2로 여름에 외부 일사량이 여름보다 높게 나타났다. 반면에 내부 일사량은 여름에는 375.7J/cm2, 겨울에는 590.7J/cm2로 겨울에 온실 내부의 일사량이 높았다. 일반적으로 외부 일사량은 겨울보다 여름에 더 높게 나타나지만, 본 연구를 위한 온실 재배 환경에서는 온실 내 온도를 낮추기 위해 외부 차광막을 함께 활용하기 때문에 여름보다 겨울에 더 높은 내부 일사량이 관측되었다.
작물의 생육은 온도와 일사량 이외에도 여러 가지 환경 요인으로부터 복합적인 영향을 받는다. 온도가 낮아지면 개화가 늦어지고 생육이 지연되며(Adams 등, 2001, Van Ploeg와 Heuvelink, 2005), 일사량에 따라 토마토의 수량 및 품질이 달라진다고 알려져 있다(Adams 등, 2001; Gent 등, 2007). 본 연구의 경우 냉난방 시설이 설치된 반밀폐형 첨단디지털온실에서 재배된 토마토의 생장을 관찰하였으며, 이러한 이유로 재배 시기와 관계없이 온실 내부 온도는 유사하였으나 일사량에서는 차이를 나타내었다. 외부 일사량은 여름보다 겨울에 높아 일반적인 재배 환경과 비슷하게 나타났지만, 온실 제어 방법으로 인해 온실 내부에서 측정된 일사량은 겨울이 여름보다 높게 나타났다. Choi 등(2008)은 일사량이 계절과 온실의 설치 방향에 따라 달라질 수 있고 이에 따른 작물의 생육 차이를 보고하였으며, Lim 등(2021)은 차광에 의한 광 환경 변화가 작물의 생육과 생산에 영향을 미친다는 결과를 보고하였다. 따라서, 본 연구의 재배 환경에서 나타난 겨울과 여름의 내부 일사량 차이가 토마토 과실이 성숙하는 과정에 영향을 미쳤을 것이라 예상해 볼 수 있다. 그러나 여기서 나타난 일사량의 차이가 토마토 과실의 성숙 속도에 미치는 유의한 영향을 확인하기 위해서는 일사량을 제외한 통제된 환경 변수를 활용한 재배를 통한 추가적인 연구가 필요할 것으로 생각된다.