

서 론
재료 및 방법
1. 실험 시설 및 환경 제어 조건
2. 실험 재료 및 광주기 처리
3. 생육 및 생리 지표 측정
4. 엽록소 형광 측정 및 OJIP 분석
5. 통계분석
결과 및 고찰
서 론
상추(Lactuca sativa L.)는 빠른 생장 속도와 짧은 재배 기간을 가지며, 인공 조명을 사용하는 식물공장 시스템에서 안정적으로 재배할 수 있는 대표적인 엽채류 작물이다. 특히 LED 조명을 활용한 식물공장 환경에서 균일한 생육과 품질 관리가 가능하여, 광환경 조건에 따른 식물 반응을 비교·분석하기 위한 모델 작물로 널리 활용되고 있다(Kelly et al. 2020; Wu et al. 2025).
한편, 광량이나 광주기와 같은 광환경 조건이 상추의 생리적 요구와 불일치할 경우 잎 생장과 형태적 품질이 변화하는 것으로 보고되고 있으며, 이러한 광환경 차이는 상추의 생육 특성에 영향을 미칠 수 있다(Dou et al. 2017).
이와 같은 인공광 환경과 달리 노지 재배 환경에서는 광량이나 광주기와 같은 광환경 조건이 식물의 생리적 요구와 일치하지 않는 경우가 빈번하게 발생하며, 이러한 조건에서는 잎 생장과 형태적 품질이 변화하는것으로 보고되고 있다. 이러한 맥락에서 Vialet-Chabrand et al.(2017) 은 시간적으로 변동하는 광 노출이 포함된 광주기 조건에서 재배된 상추에서 잎이 더 얇아지고 잎의 광 흡수율이 감소하였으며, 생육 초기 단계에서 생장 속도 또한 유의적으로 저하되었음을 보고하였다. 또한 Lawson et al.(2012)은 자연 환경과 유사한 광주기 변동 조건이 생육 기간 광합성 관련 반응의 시간적 조절에 영향을 미치며, 광주기 조절이 중요한 요인으로 작용함을 제시하였다. 이처럼 자연 환경에서는 광주기가 일정하게 유지되지 않는 경우가 많으며, 이러한 광주기 변동은 상추의 생육 및 광합성 관련 특성에 영향을 미칠 수 있음이 보고되어 왔다. 광주기는 단순한 조명 시간의 길이를 넘어, 하루 동안 식물이 경험하는 광 신호의 구조를 규정하는 환경 요인으로, 식물의 생리적 반응 전반을 조율하는 역할을 한다. 기존 연구들에서는 광주기가 탄소 동화와 에너지 이용 전략, 광합성 관련 과정의 일일 리듬 형성, 그리고 생리적 안정성 유지와 밀접하게 연관되어 있음을 제시하였으며(Osnato et al. 2022), 이에 따라 광주기는 식물 생육 반응을 해석하기 위한 핵심 환경 변수로 인식되고 있다.
또한, LED 기반 식물공장 환경에서 광의 공간적 분포 특성 역시 상추의 광합성과 생육에 중요한 영향을 미치는 것으로 보고되었으며, 산란광을 활용하여 광을 균일하게 확산시킬 경우 캐노피 내 광 이용 효율과 생장이 향상되는것으로 나타났다(Kang et al. 2016).
그러나 이러한 선행연구들은 주로 하루 총 광노출 시간의 차이 또는 연속 조명과 같은 극단적 조건에 초점을 두고 수행되어 왔으며, 동일한 총 광량을 유지하되 광과 암의 시간적 배열이 상이한 조건, 예를 들어 연속 조명과 간헐 조명 간의 비교를 통해 PSII 기능 안정성과 광화학 반응의 동역학적 조절 차이를 정량적으로 분석한 연구는 상대적으로 제한적이다. 특히 식물공장 환경에서 구현 가능한 다양한 광주기 패턴이 PSII 기능 안정성에 미치는 영향을 정량적으로 비교한 연구는 아직 충분하지 않다(Wang et al. 2021).
이러한 PSII 기능 변화와 광합성 적응 반응을 민감하게 파악하기 위한 대표적인 생리 지표로는 엽록소 형광 유도곡선인 OJIP 곡선이 널리 활용되고 있다(Sarewicz et al. 2018). OJIP 곡선은 암순화된 잎에 빛을 조사할 때 나타나는 O–J–I–P 단계의 형광 변화를 통해 PSII 전자수용체 풀의 환원 상태, 에너지 분배 양상, 그리고 광화학계의 적응 및 회복 과정을 종합적으로 반영한다(Fu et al. 2012). 이러한 특성으로 인해 엽록소 형광 분석은, 서로 다른 광주기 조건에서 PSII 기능 안정성, 전자전달 병목 현상, 그리고 광화학계의 적응 전략을 정량적으로 비교하는 데 효과적인 도구로 평가되고 있다(Kalaji et al. 2016; Küpper et al. 2019). 또한, 실제로 광 조사 직후 수 밀리초에서 수 초 범위에서 발생하는 전자전달 초기 과정이 PSII 기능 변화를 결정하는 핵심 단계로 중요성이 강조되고 있다(Kalaji et al. 2016) 그럼에도 불구하고 기존 OJIP 연구들은 주로 광강도 변화나 환경 스트레스 조건에 초점을 두어 왔으며, 동일한 일일 광적분량을 유지한 상태에서 광과 암의 시간적 배열 차이에 따른 PSII 전자전달 동역학을 체계적으로 비교한 연구는 상대적으로 제한적이다(Khan et al. 2021). 따라서 본 연구의 목적은 인공광 기반 식물공장 조건에서 서로 다른 광주기 하에 상추의 PSII 전자전달 동역학과 생육 반응 간의 관계를 OJIP 분석을 통해 정량적으로 규명하고 이를 바탕으로 합리적인 광주기 설계를 위한 생리적 근거를 제시하는데 있다.
재료 및 방법
1. 실험 시설 및 환경 제어 조건
본 연구는 경상국립대학교에 구축된 제어형 식물공장 시스템에서 수행되었다. 실험은 총 21일간 진행되었으며, 재배 기간 동안 식물공장은 온도, 상대습도, 광도 및 이산화탄소 농도를 정밀하게 제어할 수 있는 환경 제어 시스템을 갖추고 있었다. 모든 환경 변수는 고정밀 환경 센서 시스템(Hanam Engineering Co. Ltd., Hanam, Korea)을 이용하여 매일 자동으로 수집하였다.
광주기 패턴에 따라 엽 표면 온도가 단기적으로 변동할 가능성을 고려하여(Nelson and Bugbee 2014), 본 연구에서는 모든 처리구에서 공기 온도, 상대습도 및 광원을 동일하게 제어하였다. 실험 기간 동안 공기 온도는 15-20 ℃, 상대습도는 45-50 %, 이산화탄소 농도는 1,000 ± 200ppm으로 유지하였으며 광량은 250 ± 50µmol m-2 s-1 범위로 설정하였다. Amitrano et al.(2021) 에서 제시된 최적 농도 조건을 기반으로, AminoQuelant®-Ca/Mg/B(Bioiberica S.A.U., Barcelona, Spain)는 Ca, Mg, B, Mo를 포함하는 보조 영양 공급제로, 0.2%(v/v) 농도로 희석(10mL/5L)하여 각 개체에 매일 30mL씩 관주 처리하였다. 본 처리는 완전 양액이 아닌 보조 원소 공급을 목적으로 수행되었다. 상추는 비전바이오 상토 2호(Vision Bio Soil No. 2, 농우바이오, Suwon, Korea)를 이용한 배지경 조건에서 재배하였다.
2. 실험 재료 및 광주기 처리
실험 작물은 상추(Lactuca sativa L. var. crispa LIN)를 사용하였다. 종자는 파종 후 4일째부터 발아가 시작되었으며, 약 14일 후 본엽이 전개되었고 처리구 간 비교의 일관성을 확보하기 위해 모든 처리구에서 각 개체의 동일한 생리적 단계에 해당하는 본엽을 선정하여 측정을 수행하였다. 광주기 조건은 L1/D1, L6/D6, L12/D12의 세 처리구로 구성하였다. 상추 재배에서 일반적으로 활용되는 L12/D12 광주기를 기준 조건으로 설정하였으며, 이는 “Plant Factory”(2016),에서 제시된 재배 체계를 기반으로 하였다. 또한 동일한 일일 광조사 시간을 유지한 상태에서 광과 암기를 6시간 단위로 분할하여 L6/D6 처리구를 구성하였다.
L1/D1 처리구는 광과 암을 1시간 단위로 반복하여 배열함으로써 광주기의 시간적 패턴 차이를 비교할 수 있도록 설계하였다. 모든 처리구에서 하루 24시간 중 총 12시간 동안 광조사가 이루어지도록 동일한 일일 광조사량(DLI)을 유지하였다. 이러한 실험 설계는 광의 총량을 통제한 상태에서 광주기 패턴에 따른 시간적 분배 효과만을 분리하여 분석하였다. 식물 생육 특성은 개체 간 변동성이 크게 나타날 수 있으므로, 충분한 반복 수 확보가 필요하다(Poorter et al. 2012). 이에 따라 본 연구에서는 20-30개체 수준을 기준으로 각 처리구당 25개체를 배치하였다.
3. 생육 및 생리 지표 측정
본 연구에서는 식물의 생육 및 광합성 반응을 종합적으로 평가하기 위해 엽장, 엽폭, SPAD 값 및 엽록소 형광과 같은 비파괴적 지표와, 뿌리 길이, 엽면적, 생체중 및 건물중과 같은 파괴적 생육 지표를 측정하였다. 엽장, 엽폭 및 SPAD 값은 생육 상태를 대표하는 비파괴적 지표로 널리 활용되며, Lee et al.(2012) 의 측정 기준에 따라 생리적으로 성숙한 잎을 대상으로 분석하였다. 상단 본엽을 생리적 반응 평가에 활용하는 접근은 환경 스트레스에 대한 민감도가 높다는 연구결과에 따라(Sheikhi et al. 2024), 상단 본엽을 비파괴적 생육 지표의 측정 부위로 선정하였다.
이에 따라 본 연구에서는 비파괴측정을 각 개체에서 동일한 생리적 단계에 해당하는 본엽 3장을 선정하였고, 잎의 중앙 부위를 기준으로 엽장, 엽폭 및 SPAD 값을 측정하였다. 엽장은 Nakanwagi et al.(2018)의 측정 기준에 따라 잎을 세로 방향으로 두었을 때 중앙 하단에서 최상단까지의 길이를 기준으로 측정하였으며, 엽폭은 동일한 잎의 중앙부를 가로 방향으로 측정하였다. 엽장, 엽폭 및 뿌리 길이는 디지털 버니어캘리퍼스(CD-15CP, Mitutoyo, Kanagawa, Japan)를 사용하여 측정하였고, 잎 면적은 ImageJ 프로그램을 이용하여 분석하였다. 엽록소 함량은 휴대용 엽록소 측정기(SPAD-502Plus, Konica Minolta, Japan)를 사용하여 SPAD 값으로 평가하였고 뿌리 길이는 하배축을 기준으로 주근 끝단까지의 직선 거리를 디지털 버니어캘리퍼스로 측정하였다.
파괴적 생육 지표는 각 처리구에서 동일한 개체 수를 대상으로 측정하였으며, 처리구별 평균값을 산출하여 분석에 활용하였다. 반면, 비파괴적 지표는 21일간 하루 1회 반복 측정하였고, 각 개체에서 선정한 3개 잎의 측정값을 평균하여 개체별 대표값을 산출한 후, 이를 기반으로 시계열 분석을 수행하였다.
4. 엽록소 형광 측정 및 OJIP 분석
엽록소 형광은 본엽 상단부의 건전한 잎을 대상으로 완전한 암실에서 30분간 암적응을 실시한 후 엽록소형광측정장치(OS-30p+, ADC BioScientific Ltd., UK)을 이용하여 측정하였다. 장비는 3500µmol m-2 s-1의 강도로 2초간 조사되는 포화 적색광을 제공하여 OJIP 형광 유도 신호가 기록되도록 설정하였고, 각 잎은 동일한 조건에서 3회 반복 측정 후 평균값으로 분석하였다. 처리구당 총 25개의 잎을 측정하여 최종적으로 OJIP 형광 유도 반응을 측정하고 측정된 형광 곡선을 기반으로 분석되었으며, 본 연구에서 사용된 각 JIP-test 파라미터의 정의와 생리적 의미는 Table 1에 정리하였다.
Table 1
Chlorophyll fluorescence parameters and their definitions
5. 통계분석
Table1에 제시된 엽록소 형광 파라미터와 Table 2에 생육 지표의 처리 간 차이는 일원분산분석(one-way ANOVA)을 수행한 후 Duncan의 다중비교검정을 통해 분석하였다. 동일 파라미터 내에서 서로 다른 문자로 표시된 평균값은 p < 0.05 수준에서 통계적으로 유의한 차이를 나타낸다. 엽록소 형광 지표와 생육 특성 간의 상관관계는 Pearson 상관분석을 통해 평가하였다. 모든 통계 분석은 SPSS version 20.0(IBM, New York, NY, USA)을 사용하여 수행하였다. 그래프 시각화는 Origin 2024와 Microsoft Excel 2010을 이용하여 레이더 차트 및 상관 행렬 형태로 표현하였다.
Table 2
Effects of different photoperiod regimes on growth parameters of lettuce after 21 days of treatment
결과 및 고찰
광주기 조건에 따른 상추의 생육 반응을 종합적으로 평가하기 위해 초장, 뿌리 길이, 엽면적, 생체중, 건물중 및 SPAD 값을 포함한 주요 생육 지표를 분석하였다. 이러한 지표들은 상추의 형태적 생장뿐 아니라 생리적 상태와 생산성을 종합적으로 반영하는 핵심 변수들이다. 그 결과, Table 2에 제시된 바와 같이 광주기 패턴에 따라 상추의 형태적 및 생리적 생육 특성이 구분되었다. 상추의 초장과 뿌리 길이는 L1/D1 처리구에서 각각 140.10mm와 94.82mm로 가장 높은 값을 나타냈으며, 이는 짧은 광·암 반복 조건에서 상추의 초장과 뿌리의 생장이 동시에 촉진되었음을 의미한다. 생체중과 건물중 역시 L1/D1 처리구에서 각각 37.61g과 1.68g으로 가장 높았고, 반대로 L6/D6 처리구에서는 건물중이 다른 처리구에 비해 유의적으로 낮았다(p < 0.05). 엽면적 또한 L1/D1 처리구에서 가장 넓었으며, L6/D6 처리구에서 현저히 감소하였다. 한편, SPAD 값은 L12/D12 처리구에서 가장 높게 나타나, 광주기 조건이 생육량뿐 아니라 엽록소 축적에도 서로 다른 영향을 미침을 보여주었다. 이러한 결과는 광환경 조성이 상추의 생육과 광합성 관련 특성을 동시에 조절한다는 최근 연구 결과와도 일치한다(Shin and Lee 2025).
이와 같은 처리구 간 생육 및 엽록소 축적의 차이가 광합성 광화학계 수준의 기능 변화와 어떻게 연관되는지를 규명하기 위해, PSII 기능 변화를 JIP-test 기반 시계열 분석을 통해 평가하였다 Fig. 1을 보면 생육 지표에서 관찰된 처리구 간 차이의 생리적 기작을 규명하기 위해 PSII 기능 변화를 JIP-test 시계열 분석을 통해 평가하였다. 실험 개시일인 Day 1에는 모든 처리구에서 OJIP 유도 곡선의 형태가 거의 동일하게 나타나, 처리 이전 식물들이 생리적으로 균일한 상태였음을 확인하였다. Day 3 및 Day 6에서는 세 처리구 모두 O–J–I 단계까지의 형광 상승 패턴이 유사하게 유지되었으며, 이는 초기 생육 단계에서 PSII 기능이 안정적으로 유지되고 있음을 의미한다. 그러나 Day 9와 Day 12에 이르러 L6/D6 처리구에서는 J–I–P 구간의 형광 강도가 점차 감소하였고, 특히 P점 형광의 억제가 뚜렷하게 관찰되었다. 이러한 변화는 PSII 전자전달계가 충분한 포화 수준에 도달하지 못했음을 시사하며, 광·암 전환 빈도뿐 아니라 광 및 암 구간의 지속시간에 따른 에너지 축적과 회복의 불균형으로 인해 기능적 부담이 누적되었을 가능성을 보여준다.
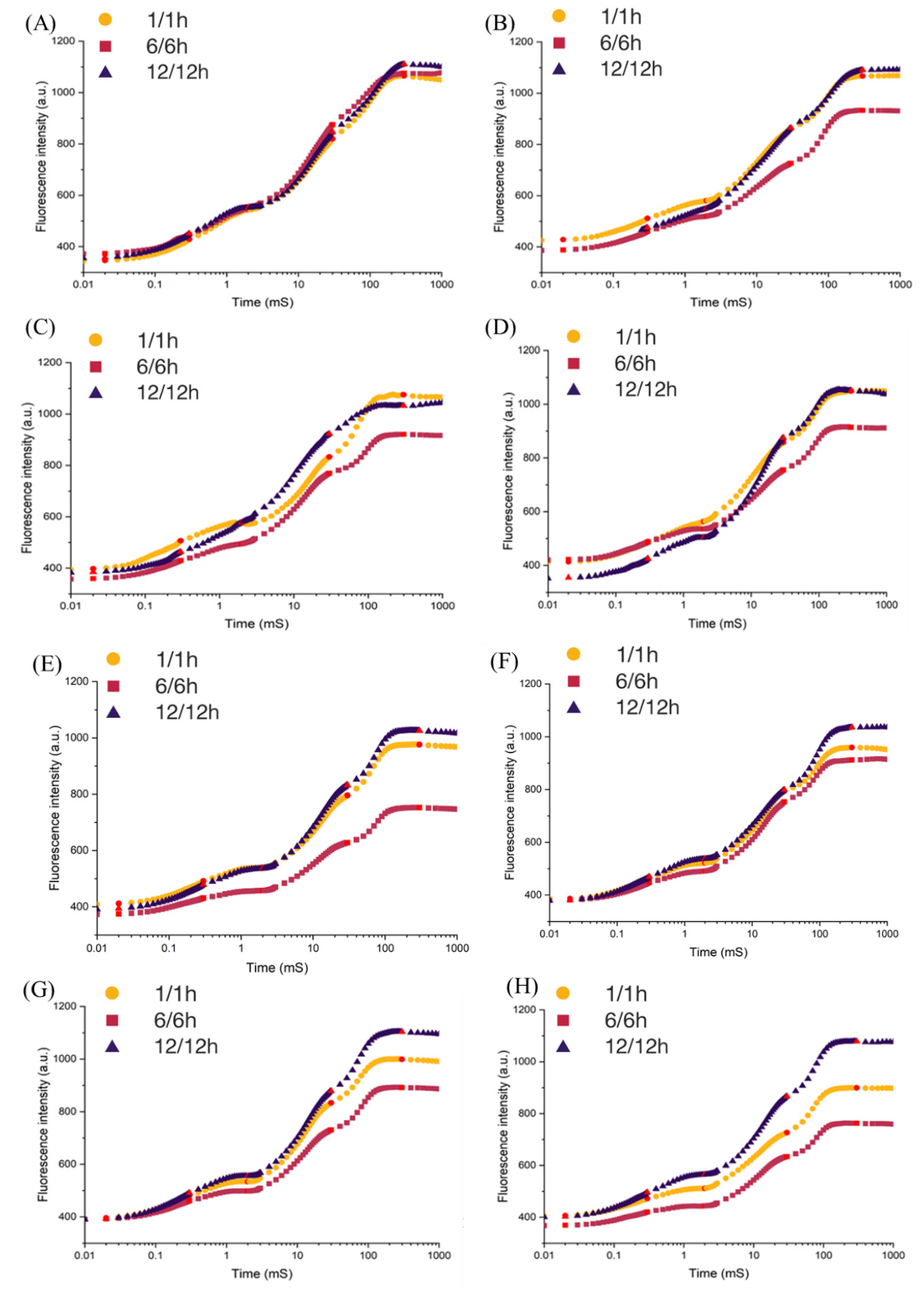
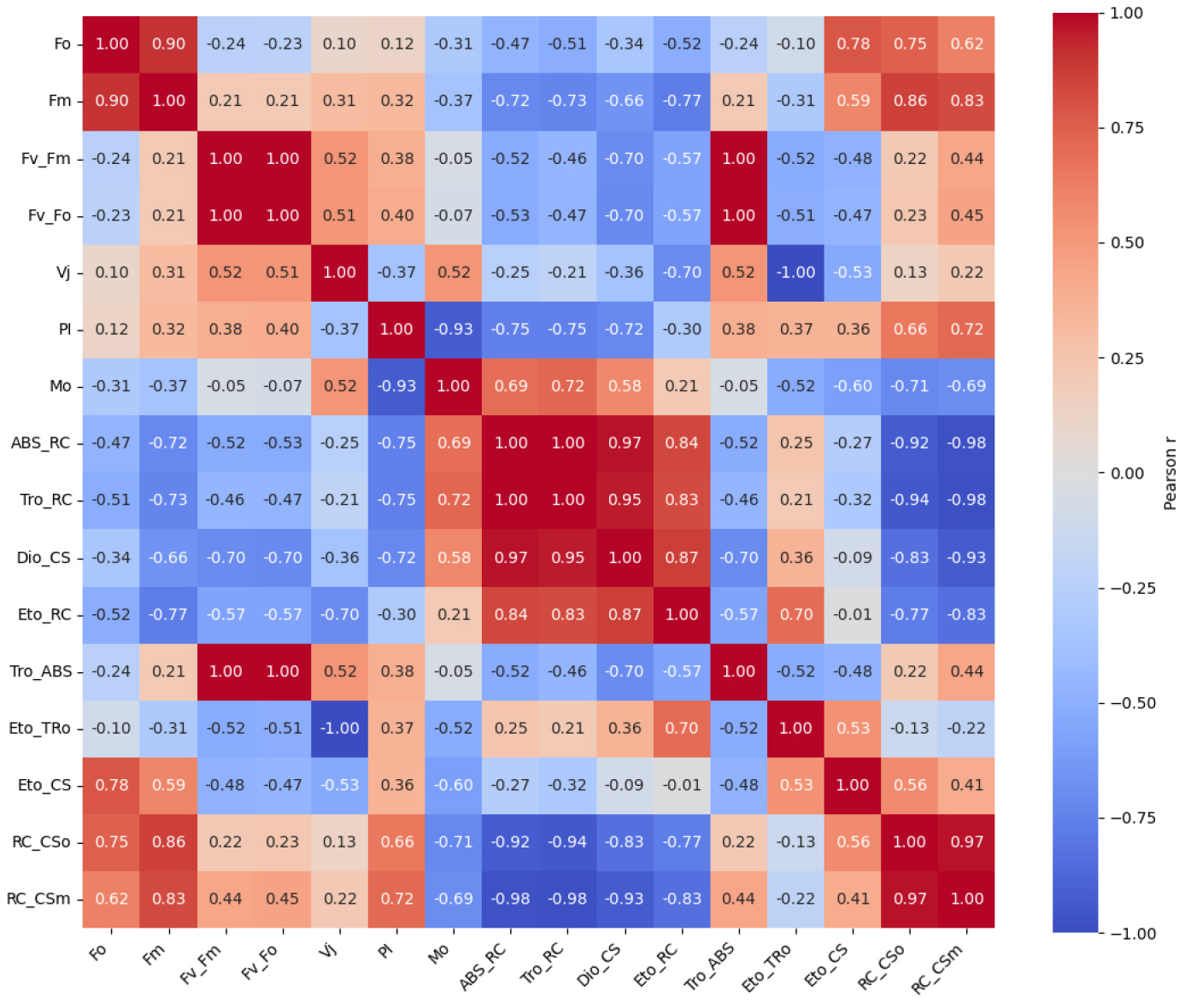
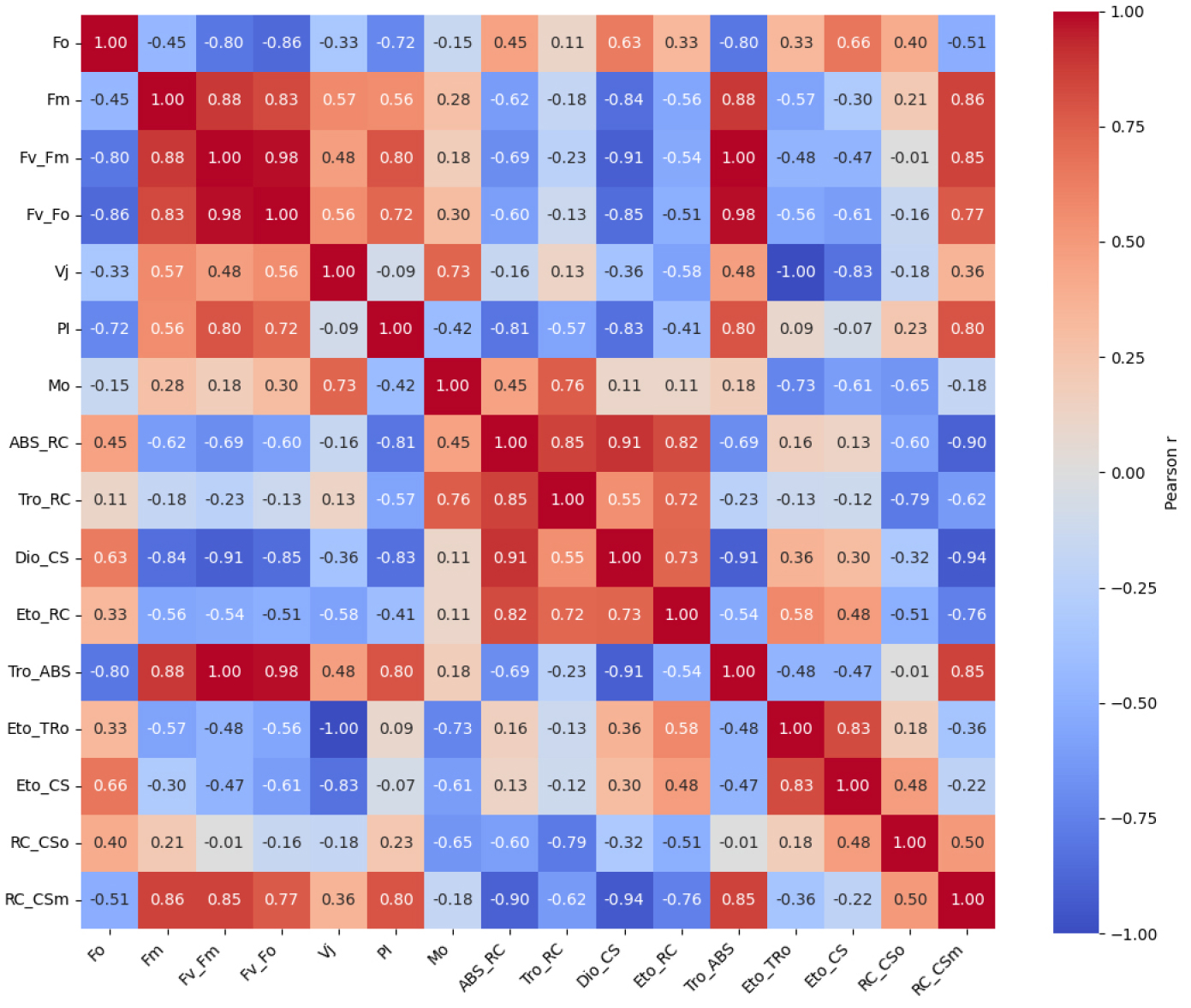
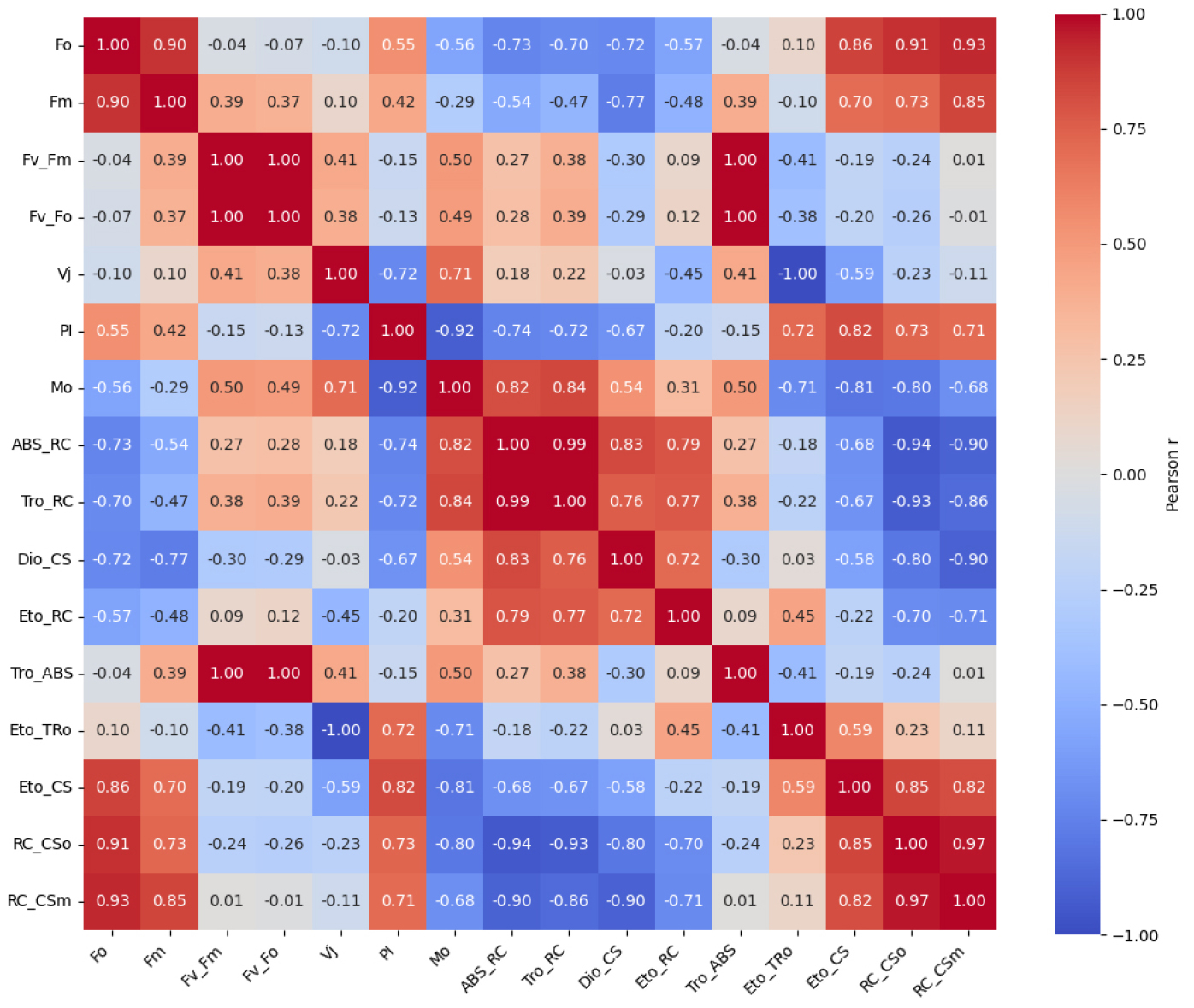
Fig. 2는 L12/D12 처리구를 기준값 1로 정규화한 PSII 파라미터의 상대적 변화를 레이더 플롯으로 나타낸 것이다. 이 분석은 광주기 조건에 따라 PSII 기능 요소들의 상대적 조합과 구조적 차이를 시각적으로 비교하기 위해 수행되었다. 전반적으로 L6/D6 처리구는 PIabs, Fv/Fm, Fv/Fo, RC/CSm, ETo/RC 등 PSII 성능을 대표하는 핵심 지표에서 일관되게 낮은 상대값을 보여, 광화학 반응 중심의 활성과 전자전달 효율이 전반적으로 저하된 구조를 나타냈다. 이는 반응중심 밀도 감소와 하류 전자전달 제한이 동시에 발생했음을 시사한다. 반면, L1/D1 처리구에서는 ABS/RC, DIo/CS, Vj, Mo와 같은 지표들이 상대적으로 증가하여, 반응중심당 에너지 부담과 초기 스트레스 반응이 강화된 상태를 반영하였다. L12/D12 처리구는 기준 조건으로서 구조적 안정성과 에너지 분배의 균형이 유지된 분포를 나타냈다. PSII 파라미터 간의 기능적 연계를 정량적으로 분석하기 위해, Figs. 3, 4, 5를 보면 각 광주기 조건별로 JIP-test 파라미터 기반 상관행렬을 도출하였다. 이러한 분석은 개별 지표의 변화뿐 아니라, 지표 간 상호작용 구조를 통해 PSII 조절 메커니즘을 해석하는 데 목적이 있다. Fig. 4를 보면 L1/D1 조건에서는 Fo와 Fm간 높은 양의 상관(r = 0.90)이 유지되었으며, Vj와 Mo 도 양의 상관(r = 0.52)을 나타냈다. 특히 Vj-PI(r = -0.37)와 Mo-PI(r = -0.93)의 음의 상관은 전자전달 초기 병목이 PSII 전체 성능 저하를 결정하는 주요 요인임을 보여준다. Fig. 4 L6/D6 조건에서는 ABS/RC, DIo/CS와 PI 간의 강한 음의 상관과 함께, ABS/RC와 RC/CSm 간의 매우 강한 음의 상관(r = -0.90)이 관찰되어, 반응중심 밀도 감소와 에너지 부담 증가가 직접적으로 연결됨을 시사하였다. 반면 L12/D12 조건에서는 구조적 지표와 전자전달 효율 간의 균형적 상관 구조가 유지되었다.
이상의 결과를 종합하면, 분절된 광주기 조건인 L6/D6에서는 PSII 반응중심의 손상과 회복 간 시간적 균형이 유지되지 못하면서 기능 저하가 누적된 것으로 해석된다. D1 단백질 손상은 비교적 짧은 시간 규모에서 발생하는 반면, 분해와 신규 합성을 포함하는 복구 과정은 더 긴 시간 규모를 요구하는 것으로 알려져 있다(Baker 2008). 이러한 상황에서 반복적인 광·암 전환은 손상된 PSII가 충분히 회복되기 전에 다시 광에 노출되는 조건을 형성하여, 기능 저하를 가속했을 가능성이 있다(Didaran et al. 2024). 반면, Fig. 5에서 나타난 L12/D12 조건에서의 PSII 안정성 유지는 충분한 암기 회복 시간이 확보된 결과로 볼 수 있다. 그러나 이러한 결과는 단순한 암기의 절대적 지속시간보다는 명기와 암기의 상대적인 비율이 전자전달계의 에너지 축적과 회복 간 균형을 유지하는 데 더 큰 영향을 미쳤기 때문으로 해석된다(Murata et al. 2007).
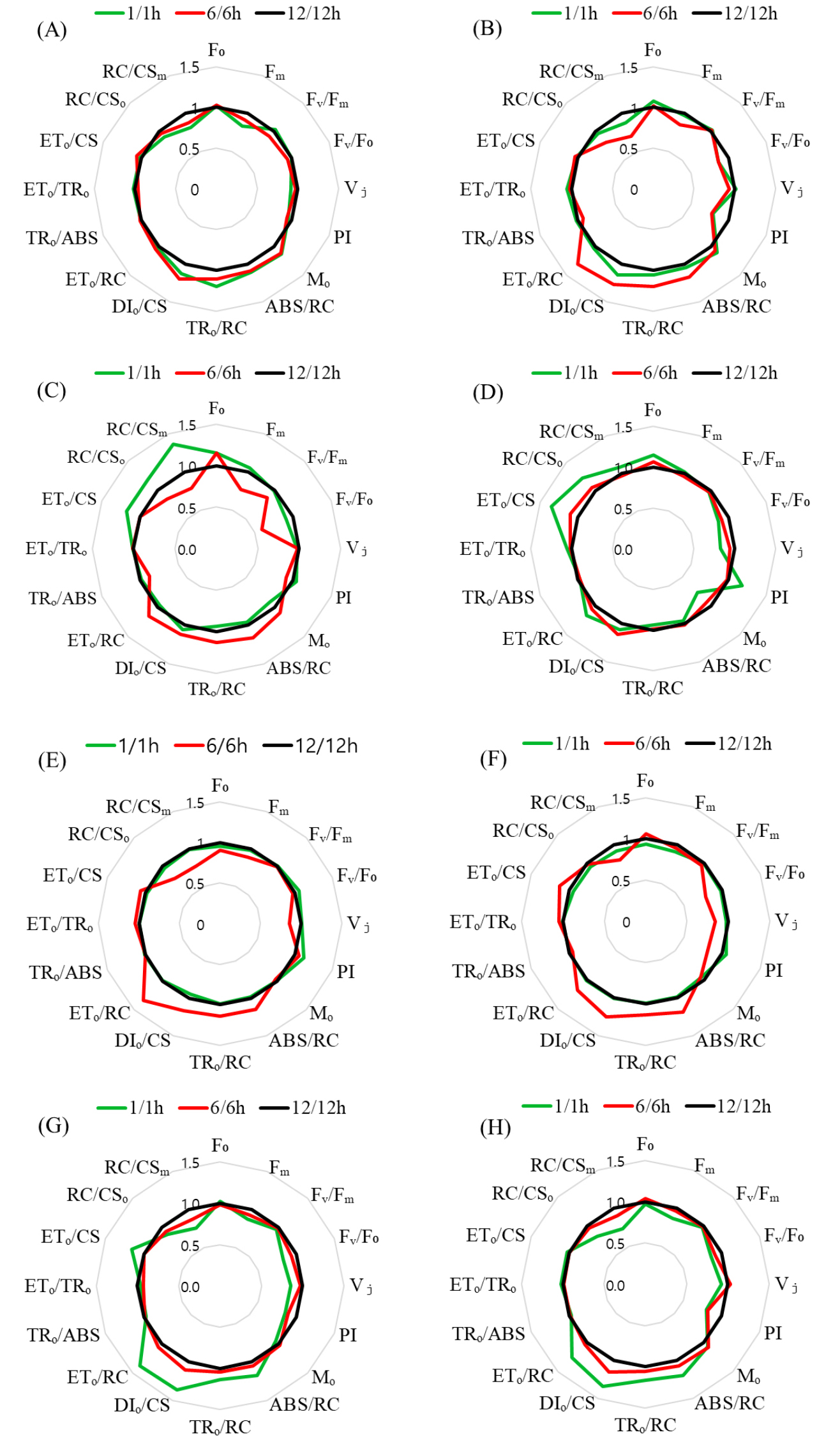
Fig. 2
Radar plot analysis of chlorophyll a fluorescence induction (OJIP) parameter in lettuce grown under three photoperiod treatments (L1/D1, L6/D6, L12/D12). The spider plots illustrate major OJIP-derived parameters, including the initial fluorescence (Fo), maximum fluorescence (Fm), maximum quantum yield of PSII (Fv/Fm), relative variable fluorescence at 2 ms (Vj), relative variable fluorescence at 30 ms (Vi), performance index on an absorption basis (PIabs), specific energy fluxes per reaction center (ABS/RC, TRo/RC, ETo/RC, DIo/RC), and electron transport–related parameters (ETo/TRo, ETo/CS, RC/CSm). Measurements were conducted on (A) Day 3 (B) Day 6 (C) Day 9 (D) Day 12 (E) Day 13 (F) Day 16 (G) Day 19 (H) Day 21. All values were normalized to the 12/12h photoperiod to visualize relative deviations across treatments and parameters (Continued)
이는 Yamori(2016)의 연구에서 보고된 PSII 구조적 안정성 유지 결과와 같은 맥락으로 해석된다. L1/D1 조건에서도 짧은 암기를 통한 빠른 회복이 가능했던 것으로 보이며, 이는(Khan et al. 2025)의 단주기 광주기 조건에서의 PSII 기능 유지 결과와 유사한 경향을 보였다.
본 연구는 서로 다른 광주기 조건에서 상추의 생육 반응과 PSII 전자전달 특성 간의 관계를 생육 지표와 OJIP 파라미터를 통합하여 단계적으로 분석하였다. 그 결과, 광주기 길이는 단순한 일일 광량의 차이가 아니라, PSII 기능의 안정성과 회복 동역학을 조절하는 주요 환경 요인임을 확인하였다. 특히 광주기 구성에 따라 전자전달 초기 병목, 반응중심 밀도, 에너지 분배 특성이 차별적으로 조절되며, 이러한 차이가 최종적인 생육 성과로 연결됨을 정량적으로 제시하였다. 이러한 결과는 인공광 기반 재배 시스템에서 광주기 설계 시 생육량뿐 아니라 생리적 안정성을 함께 고려해야 함을 시사하며, 광환경 최적화를 위한 과학적 근거를 제공한다.
