

서 론
재료 및 방법
1. 실험재료
2. 식물호르몬 분석
3. 통계 분석
결 과
1. 과실의 성숙단계에 따른 크기 변화
2. 과실의 성숙단계에 따른 IAA 함량 변화
3. 과실의 성숙단계에 따른 지베렐린 함량 변화
4. 과실의 성숙단계에 따른 지아틴 함량 변화
5. 과실의 성숙단계에 따른 앱시스산 함량 변화
6. 과실의 성숙단계에 따른 자스몬산 함량 변화
7. 과실의 성숙단계에 따른 살리실산 함량 변화
고 찰
서 론
딸기(Fragaria ananassa Duch.)는 장미과에 속하는 초본성 다년생 작물로서 전 세계적으로 배수성이 다양한 24개의 야생종이 분포하고 있는데, 현재 우리나라에서 재배되고 있는 딸기는 향기와 과색이 우수한 버지니아 종(F. virginiana)과 과실의 크기가 큰 칠레종(F. chiloensis)의 자연교잡종으로 8배체 작물이다(Liston 등, 2014). 딸기는 우리나라의 시설 채소작물 중에서 생육적온이 가장 낮은 저온성 작물로 난방비 부담이 적어 저온기 시설재배에 유리한 작물이다. 우리나라의 딸기 재배역사는 비교적 짧은 편이지만, 국내 환경에 적합한 재배작형이 잘 확립되어 있고 우수한 품종이 육성 보급되어 고품질의 신선 딸기가 생산되고 있다. 특히 딸기는 다른 시설작물에 비해 가격이 안정화되어 시설재배 농가의 주요 소득작물로 자리 잡고 있는데, 2022년도 국내 딸기의 재배면적과 생산량은 각각 5,745ha와 158,807톤이다(MAFRA, 2022).
딸기와 같은 과실의 성숙은 수량, 품질, 생리활성물질 및 호르몬 변화 등에 영향을 미치는 복잡한 대사과정을 동반한다(Anderews, 1995; Rashida 등, 1997; Li 등, 2019). 과실의 성숙과정은 유전적 또는 환경적 요인으로 생리적 또는 생화학적 변화가 일어나는데(Cherian 등, 2014; Fenn과 Giovannoni, 2021), 이러한 변화는 일반적으로 엽록소의 함량 감소, 안토시아닌이나 카로티노이드의 축적, 세포벽의 변형에 의한 질감 변화뿐만 아니라 유기산, 당, 생리활성물질 등과 같은 대사물질의 증가로 인한 품질 변화 등을 포함한다(Alvarez-Fernandez 등, 2014; Kumar 등, 2014; Giovannoni 등, 2017). 과실의 성숙 과정에 일어나는 대표적인 생리적 변화에는 가용성 당의 함량 변화(Jeong 등, 2016), 세포벽 분해와 연화 현상(Brady 등, 1985), 과실 색의 변화(Carrari와 Fernie, 2006), 생리활성물질의 축적(Ko 등, 2017) 및 호르몬의 변화(Farcuh 등, 2019; Han 등, 2019; Perez-Llorca 등, 2019) 등으로 아주 다양하다. 특히 식물호르몬은 생체 내에서 생합성되는 신호전달 물질로 아주 적은 양으로 식물의 생장과 발육 및 여러 가지 생리적 반응을 조절한다(Taiz 등, 2015). 생리적 반응 등을 기준으로 식물호르몬은 옥신, 지베렐린, 시토키닌, 에틸렌, 앱시스산으로 구분하고 있는데, 이러한 호르몬은 생장 촉진제와 생장 억제제로 분류하기도 한다(Guleria 등, 2021). 생장을 촉진시키는 호르몬으로는 옥신, 지베렐린 및 시토키닌 등이 포함되며, 세포 분열과 생장, 개화 결실 및 성숙 등을 통하여 생장을 촉진시킨다. 생장을 억제시키는 호르몬으로는 에틸렌과 아브시스산으로 휴면, 노화 및 탈리 촉진 등을 통하여 생장과 발육을 억제시킨다. 또한 자스몬산, 살리실산, 브라시노스테로이드 및 스트리고락톤 등도 새로운 식물호르몬으로 간주하기도 한다(Symons 등, 2006; Zaharah 등, 2012; Taiz 등, 2015). 과실의 성숙과정에 동반되는 호흡이나 호르몬의 급격한 변화에 따라 과실은 호흡 급등형과 비급등형으로 구분되는데, 호흡 급등형 과실에 비하여 비급등형 과실의 식물호르몬 변화는 잘 알려져 있지 않다(Adams-Phillips 등, 2004; Symons 등, 2012). 딸기는 성숙과정에 에틸렌의 생성이나 호흡은 크게 변화되지 않으며(Frenkel, 1972) 성숙은 에틸렌의 활성에 의해서 크게 유도되지 않는다고 보고하였다(Oh 등, 2018). 최근에 앱시스산이 비급등형 과실의 성숙과정에 중요한 역할을 한다고 보고되었는데(Beril 등, 2011; Shen 등, 2014; Oh, 2017), 앱시스산은 딸기 과실의 연화와 더불어 착색을 촉진시킨다고 하였다(Jia 등, 2011; Oh 등, 2018). 반면에 옥신은 비급등형 과실에 있어서 과실의 성숙을 억제시키는 역할을 하는데(Trainotti 등, 2005), 딸기 과실에서 옥신의 함량은 개화와 미성숙 과실의 비대기에 가장 높았지만 변색기 이후로는 감소한다고 보고도 있다(Zhang 등, 2003a). 또한 과실의 외부에 붙어 있는 종자에 의해 생성된 옥신은 성숙을 지연시키는 역할을 하기도 한다(Given 등, 1988; Symons 등, 2012). 비급등형 과실인 딸기에 있어서 옥신과 앱시스산은 과실의 발육에서 성숙으로 전환되는 과정에 반응하는 주된 호르몬으로 알려져 있는데, 딸기에서 옥신은 화탁의 발육과 동시에 성숙 관련 유전자의 발현을 억제시켜 성숙을 지연시킨다(Fuentes 등, 2019). 앱시스산은 성숙과 관련된 많은 유전자의 발현을 조절하지만, 에틸렌이나 지베렐린은 성숙과정 중에는 큰 역할을 하지 못하는 것으로 알려져 있다. 한편 지베렐린과 옥신에 의해서 시작되는 착과는 자방이 과실로 발육하는 과정으로 세포분열과정이 포함되는데, 착과된 과실에 있어서 세포의 팽창은 지베렐린에 의해서 일어나며 세포의 생장은 옥신과 지베렐린의 양이나 신호전달에 크게 좌우된다고 알려져 있다(Shinozaki 등, 2020). 또한 지베렐린은 딸기 과실의 성숙을 지연시킨다고 하였으며(Mukkun와 Singh, 2009), 시토키닌은 꽃눈분화와 착화수 및 착과에 영향을 미친다고 하였다(Yamasaki와 Yamashita, 1993). 딸기에 있어서 지아틴 함량은 개화와 더불어 급격하게 증가하다가 그 이후로는 감소한다는 보고도 있다(Eshghi와 Tafazoli, 2007). 최근에 식물계에 광범위하게 분포하는 리로레산에서 유기되는 자스몬산은 새로운 식물생장조절물질로 간주되고 있다(Jin과 Zhu, 2017). 자스몬산은 식물체의 생장 억제, 노화촉진, 발아와 꽃눈분화 억제, 기공 폐쇄 촉진 및 과실의 성숙촉진 등 다양한 생리적 기능이 보고되었다(Chou와 Kao, 1992; Meir 등, 1996; Herde 등, 1997; Khan과 Singh, 2015). 또한 새로운 호르몬으로 간주되고 있는 살리실산은 식물체내에서 발견되는 페놀화합물로 병원균에 대한 획득저항성에 관여한다고 한다(Symons 등, 2006; Babalar 등, 2007; Wang 등, 2020). Perez-Llorca 등(2019)은 과실의 발육과 성숙과정에 있어서 살리실산은 복합적인 역할을 한다고 하였는데, 특히 수확 전후의 생물학적 스트레스에 대한 방어 시스템의 발현에 관련된다고 하였다(Zhang 등, 2003b). 일반적으로 살리실산의 함량은 과실의 발육과정에는 높은 함량을 보이다가 성숙과 더불어 감소한다고 하였다(Oikawa 등, 2015). Shi와 Zhang(2012)은 살리실산은 ACC oxidase의 발현 억제를 통하여 에틸렌과 상호작용한다고 하여 살리실산에 의하여 에틸렌의 생성이 조절된다고 하였다.
딸기는 수정 후 과실이 비대하여 성숙되면서 점차 빨간색으로 변하는데, 품종에 따라 다양한 과실색의 변이를 나타내며 과실색은 과실의 품질을 결정하는 중요한 요소 중의 하나이다. 이러한 과실색은 안토시아닌 함량이나 조성에 의해 결정되며, 과실이 성숙하여 착색이 진행될수록 안토시아닌 함량은 급격하게 증가하는 것으로 알려져 있다(Wang과 Lin, 2000). 그러나 최근에는 성숙과정 중에 안토시아닌 색소의 합성이 줄어들거나 합성되지 않아 과피색이 연한 분홍색이거나 흰색을 나타내는 딸기가 육성되어 고급화된 특성화 품종으로 소비자의 관심을 받고 있다. 또한 딸기의 소비패턴도 대과 위주의 생식용에서 케익 등의 장식용으로 크기가 균일한 중·소과의 수요도 급증하고 있다. 따라서 본 실험은 과피색이 분홍색인 딸기와 크기가 균일한 중·소과의 생산이나 신품종 개발을 위한 기초정보를 제공하고자 수행하였다.
재료 및 방법
1. 실험재료
실험에 사용한 딸기(Fragaria ananassa Duch.) 품종은 과실의 크기가 크며 완숙과의 과실색이 적색인 설향과 분홍색인 만년설 및 과실의 크기가 작고 분홍색인 파인베리 과실을 사용하였다. 과실의 성숙단계는 수분 후 10일경인 미숙기(green), 20일경인 변색기(breaker), 30일경인 채색기(turning) 및 40일경인 완숙기(mature)로 구분하여 수행하였다(Fig. 1).

실험용 딸기는 경남 산청군 단성면에 위치한 양액재배 농가에서 2화방 과실을 착과 후 성숙단계별로 2월 10일에서 3월 20일 사이에 채취하였다. 분석용 딸기 과실은 전체 하우스에서 완전임의배치 3반복으로 반복당 3지점(9지점)을 선정한 후 각 지점에서 9개의 과실을 성숙단계별로 채취하여 과실 무게를 측정한 후 식물호르몬 분석용으로 사용하였다.
2. 식물호르몬 분석
2.1 분석 시료 준비
분석용 딸기 과실은 채취하여 초저온 냉동고(-70°C)에서 급냉시킨 후 동결건조기(Hypercool HC 3110, Hanil Scientific Inc., Korea)에서 동결건조 시켰다. 동결 건조된 딸기 시료는 균질기(Polytron, PT-MR 3100D, Kinematica, Switzerland)로 마쇄시켜 Pan 등(2010)의 방법에 준하여 분석하였다. 균일하게 마쇄된 시료 50mg을 2mL 원심분리용 튜브에 넣고 500μL 추출 용매(2-propanol/H2O/concentrated HCl; 2:1:0.002; v/v/v)를 첨가하여 4°C에서 30분간 100rpm으로 진탕시켰다. 진탕 후 1mL dichloromethan을 각 시료에 첨가하여 4°C에서 30분간 100rpm으로 다시 진탕시킨 후 4°C에서 5분간 13,000g으로 원심분리시켰다. 원심분리 후 층 분리가 일어나면 상징액 900μL을 원심분리용 새로운 2mL 튜브에 옮겼다. 회수한 상징액은 회전농축기(Hanil HyperVAC VC2124)를 이용하여 증발시키고 회수된 시료는 최종적으로 100μL 메탄올에 용해하여 분리용 시료로 이용하였다. 시료는 분석 전까지 -20°C에 보관하였다.
2.2 표준 호르몬
호르몬 분석을 위한 표준 저장용액은 표준물질로 옥신(indole-3-acetic acid, IAA), 지베렐린(gibberellic acid, GA3), 시토키닌(zeatin), 앱시스산(2-cis,4-trans-abscisic acid, ABA), 자스몬산(jasmonic acid, JA), 및 살리실산(salicylic acid, SA)을 사용하였으며, 표준 저장용액은 메탄올 1mL에 표준물질을 용해시켜 1mg/mL의 농도로 준비하였다. 내부 표준물질(IS)은 D-JA를 사용하였으며, 메탄올에 용해시켜 1mg/mL의 농도로 분석에 사용하였다.
2.3 HPLC 운용 조건
HPLC는 degasser, pump, autosampler and column oven이 장착된 Agilient 1260 series (Agilent Technologies, Palo Alto, CA, USA) 시스템으로 사용하였다. 크로마토그램 분리는 Esclipe plus C18 (4.6 × 50mm, 3.5μm) column (Agilent, USA)을 이용하였다. HPLC 이동상은 0.1% formic acid in Water과 0.1% formic acid in Methanol를 이용하였다. 기울기 조건은 처음은 5%에 시작하였으며 1분간 95%로 변화시키고 4분 동안 95%로 유지한 후 0.1분간 빠르게 5%로 변화시키고 0.9분간 유지 시켜주었다. 용매의 유속과 컬럼 오븐의 온도는 각각 0.5μL/min과 30°C로 설정하였으며, 각 시료의 주입량은 10μL로 하였다.
2.4 Mass spectrometry 운용조건
질량분석기는 ESI (electro spray ionization) source가 장착된 API 4000 mass spectrometer (SCIEX, USA) 질량분석기를 이용하였다. Negative 모드와 MRM 모드로 분석을 수행하였다. BioAnalystTM (version 1.6.1)과 analyst software (version 1.6.1) 프로그램을 각각 장비 운용과 테이터 분석에 이용하였다. 이온화 시 고순도 질소 가스를 분무하여 건조 가스로 사용하였는데, 가스의 압력은 60psi로 각각 설정하였다. 이온 분무 전압은 -4.5kV로 하였고 600°C의 이온화 소스 온도를 이용하였다. Q1과 Q3는 unit resolution을 이용하여 LC- MS/MS의 다중성분분석법(multiple reaction monitoring)으로 분석하였다.
2.5 직선성
검량곡선을 작성하기 위해 각 표준 저장용액(1mg/mL)을 이용하여 메탄올에 희석하여 사용하였는데, IAA는 5, 10, 20, 50, 100, 500, 1,000, 10,000ng/mL, GA3는 0.5, 1.0, 2.0, 5.0, 10, 50, 100, 1,000ng/mL, zeatin은 1.5, 3.0, 6.0, 15, 30, 150, 300, 3,000ng/mL, ABA와 JA는 0.15, 0.3, 0.6, 1.5, 3.0 15, 30, 300ng/mL, SA는 0.05, 0.1, 0.2, 0.5 1.0, 5.0, 10, 100 ng/mL으로 용액을 조제하였다. 위와 같이 조제한 각 표준용액을 LC-MS/MS에 주입하여 크로마토그램상에서 나타난 피크면적을 기준으로 표준선을 작성하고 상기분석 과정을 수행하여 표준용액 및 분석시료를 각각 3반복 처리 후 식물호르몬을 정량 분석하였다. 직선성은 모든 표준물질에서 r2 = 0.995 이상의 회귀성을 보여주었다.
2.6 검출한계 및 정량한계
표준 호르몬의 검출한계(Limit of Dection, LOD)와 정량한계(Limit of Quantitation, LOQ)는 LC-MS/MS 크로마토그램상에서 나타난 각 표준시료의 신호대 잡음비(S/N ratio)가 3과 10인 농도로 각각 산출하였고 그 값은 다음과 같다. IAA 3.0ng/L, GA3 0.30ng/L, zeatin 0.90ng/L, ABA와 JA 0.03 ng/mL, SA는 0.03ng/L으로 각각 검출한계(LOD)가 확인되었다. 정량한계(LOD)는 IAA 10ng/mL, GA3 1.0ng/L, zeatin 3.0ng/mL, ABA와 JA 0.3ng/mL, SA는 0.1ng/mL으로 확인되었다.
3. 통계 분석
성숙단계별로 채취된 시료의 식물호르몬 분석은 3반복으로 실시하였다. 본 실험에서 분석한 결과 값은 SigmaPlot 12.0 (SYSTAT, Chicago, IL, USA) 프로그램을 이용하여 평균과 표준편차를 계산하였고, SAS 통계 프로그램(SAS, 9.4, Institute Inc., USA)을 이용하여 Duncan’s multiple range test로 95% 유의수준에서 분석하였다.
결 과
1. 과실의 성숙단계에 따른 크기 변화
딸기 과실의 성숙단계에 따른 과실무게의 변화 양상은 전반적으로 모든 품종에서 착과와 더불어 채색기까지는 증가하는 경향을 보였지만, 품종간 차이가 뚜렷하여 설향과 만년설에 비해 파인베리 품종은 아주 작았다(Fig. 2). 설향의 미숙기 과실무게는 6.9g에서 변색기에는 15.5g으로 2.2배 증가하였고 과색이 빨간색으로 변하는 채색기에는 20.5g으로 3.0배 증가하였지만, 완숙기의 과중은 20.2g으로 채색기와 비슷하였다. 만년설의 미숙기 과실무게는 6.9g에서 과색이 변색기에는 16.6g으로 2.4배 증가하였고 과색이 분홍색으로 변하는 채색기에는 21.9g으로 3.2배 증가하였지만, 완숙기에는 23.0g으로 채색기와 비슷한 경향을 보였다. 파인베리의 미숙기 과실무게는 0.6g에서 변색기에는 1.6g으로 2.7배 증가하였고, 과색이 분홍색으로 변하는 채색기에는 3.9g으로 6.5배 증가하였으며 완숙기에는 4.6g으로 채색기와는 큰 차이가 없었다. 완숙기를 기준으로 과실의 크기는 파인베리의 4.6g에 비해 설향은 20.2g으로 4.4배, 만년설은 23.0g으로 5.0배 컸고, 과실색은 설향이 적색인데 비해 만년설과 파인베리는 분홍색이었다.

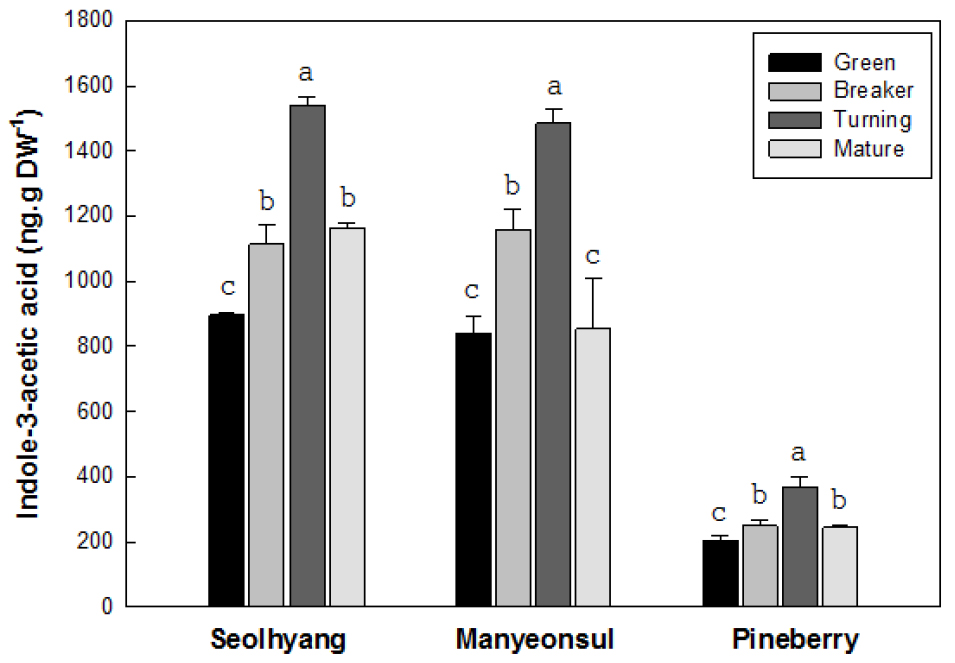
2. 과실의 성숙단계에 따른 IAA 함량 변화
딸기 과실의 성숙단계에 따른 IAA의 함량 변화는 성숙과 더불어 채색기까지는 증가한 후 완숙기에는 다소 감소하는 경향을 보였는데, 과실의 크기가 다른 품종간 양적인 차이는 뚜렷하였다(Fig. 3). 설향의 미숙기 IAA 함량은 건물 1g 당 896ng에서 과실의 크기가 가장 컸던 채색기에는 1,542ng으로 1.7배까지 증가한 후 완숙기에는 1,165ng으로 채색기에 비해 25% 감소하였다. 만년설의 미숙기 IAA 함량은 838ng인데 비해 채색기에는 1,487ng으로 1.8배 증가한 후 완숙기에는 854ng으로 채색기에 비해 43%가 감소하였다. 파인베리의 미숙기 IAA 함량은 263ng인데 비해 채색기에는 348ng으로 1.3배 증가하였고 완숙기에는 237ng으로 채색기에 비해 32%가 감소하는 경향을 보였다. IAA의 함량이 가장 높았던 채색기에서 파인베리 과실의 348ng에 비해 설향은 1,542ng으로 4.4배, 만년설은 1,487ng으로 4.3배 수준으로 높은 경향을 보여 과실의 크기에 따른 IAA 함량은 뚜렷한 차이를 보였지만 과색에 따른 차이는 없는 것으로 판단되었다.
3. 과실의 성숙단계에 따른 지베렐린 함량 변화
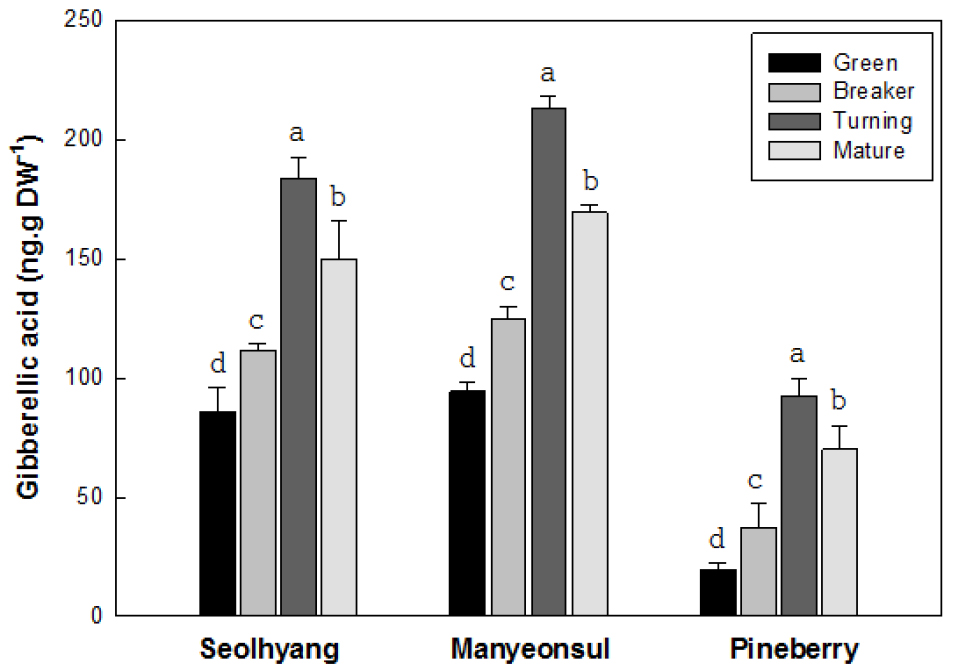
딸기 과실의 성숙단계에 따른 지베렐린의 함량은 성숙과 더불어 채색기까지는 증가한 후 완숙기에는 감소하는 경향을 보였는데, 과실 크기에 따른 품종간 차이는 뚜렷하였다(Fig. 4). 설향의 미숙기 지베렐린 함량은 건물 1g 당 85.9ng인데 비해 변색기와 채색기에는 각각 111.6과 183.4ng으로 1.3과 2.1배 증가하였고 완숙기에는 149.8ng으로 채색기에 비해 18.3% 감소하였다. 만년설의 미숙기 지베렐린 함량은 94.4ng인데 비해 변색기와 채색기에는 각각 124.9와 213.3ng으로 1.3과 2.3배 증가하였고 완숙기에는 169.3ng으로 채색기에 비해 20.6%가 감소하였다. 파인베리의 지베렐린 함량은 전반적으로 설향이나 만년설에 비해 낮은 경향을 보였는데, 미숙기의 지베렐린 함량은 19.7ng인데 비해 변색기와 채색기에는 각각 37.2와 92.2ng으로 1.9와 4.7배 증가하였고 완숙기에는 69.8ng으로 채색기에 비해 24.3%가 감소하였다. 지베렐린의 함량은 모든 품종이 채색기에서 가장 높았는데, 파인베리의 92.2ng에 비해 설향과 만년설에서는 각각 183.4와 213.3ng으로 1.9와 2.3배 수준으로 높은 경향을 보여 보여 과실의 크기에 따른 지베렐린의 함량은 뚜렷한 차이를 보였지만 과색에 따른 차이는 없는 것으로 판단되었다.
4. 과실의 성숙단계에 따른 지아틴 함량 변화
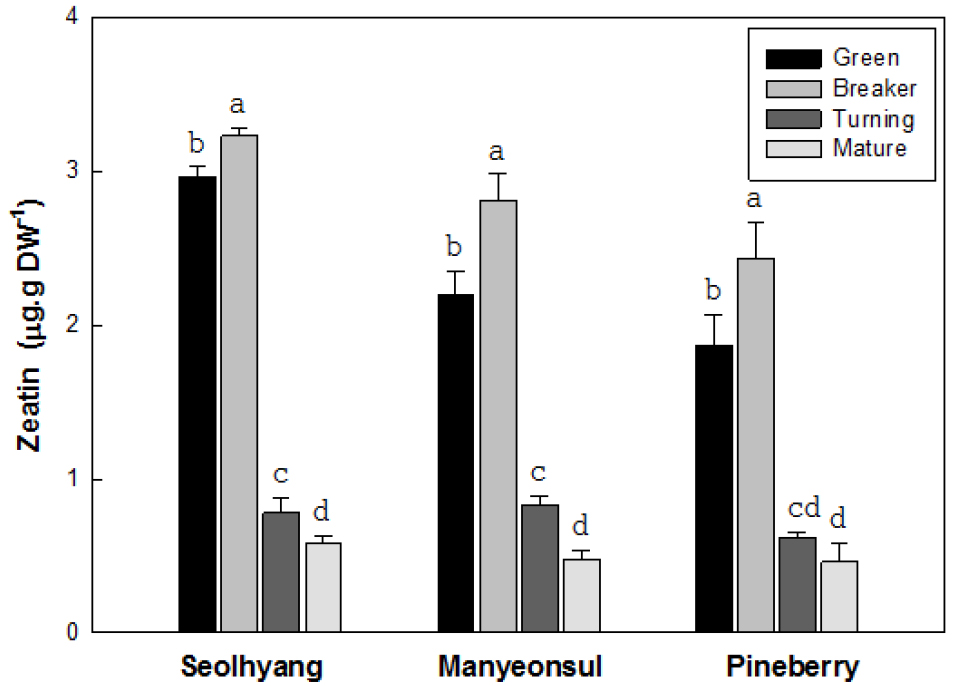
딸기 과실의 성숙단계에 따른 지아틴 함량은 모든 품종에서 미숙기에서 변색기까지는 다소 증가하다가 채색기부터는 급격하게 감소하는 경향을 보였다(Fig. 5). 설향의 미숙기 지아틴 함량은 건물 1g 당 2.9µg에서 변색기에는 3.2µg으로 다소 증가하였지만, 채색기와 완숙기에는 각각 0.8과 0.6µg으로 급격하게 감소하였다. 만년설의 미숙기 지아틴 함량은 2.2µg에서 변색기에는 2.8µg으로 증가하였지만, 채색기와 완숙기에는 각각 0.8과 0.5µg으로 급격하게 감소하였다. 파인베리의 미숙기 지아틴 함량은 1.8µg에서 변색기에는 2.4µg에서 증가한 후 채색기와 완숙기에는 각각 0.6와 0.5µg으로 급격하게 감소하였다. 그러나 과실의 크기나 과실색에 따른 지아틴 함량은 전반적으로 큰 차이가 없어 IAA와 지베렐린의 결과와는 다른 경향을 보였다.
5. 과실의 성숙단계에 따른 앱시스산 함량 변화
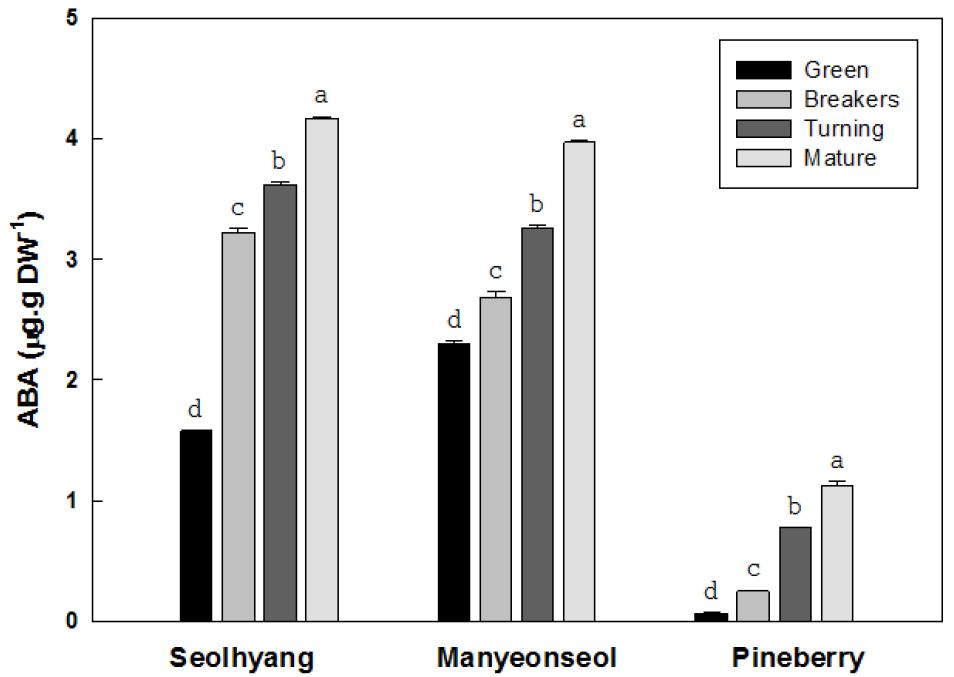
딸기 과실의 성숙단계에 따른 앱시스산의 함량은 성숙과정이 진행될수록 증가하는 경향을 보였으며, 과실 크기가 다른 품종간 차이가 뚜렷하여 과실이 큰 설향과 만년설에 비해 아주 작은 파인베리에서는 아주 낮은 경향을 보였다(Fig. 6). 설향의 미숙기 앱시스산 함량은 건물 1g 당 1.58µg에서 증가하여 변색기와 채색기에는 각각 3.22와 3.62µg으로 2.0와 2.3배 증가하였고 완숙기에는 4.13µg으로 2.6배 증가하였다. 만년설의 미숙기 앱시스산 함량은 2.30µg에서 증가하여 변색기와 채색기에는 각각 2.68과 3.22µg으로 증가하였고 완숙기에는 3.96µg으로 미숙기 대비 1.7배 증가하였다. 파인베리의 앱시스산 함량은 설향이나 만년설에 비해 낮은 경향을 보였는데, 미숙기의 앱시스산 함량은 0.07µg에서 변색기와 채색기에는 각각 0.25와 0.78µg으로 3.6과 11.1배 증가하였고 완숙기에는 1.13µg으로 미숙기 대비 16.1배 증가하였다.
6. 과실의 성숙단계에 따른 자스몬산 함량 변화
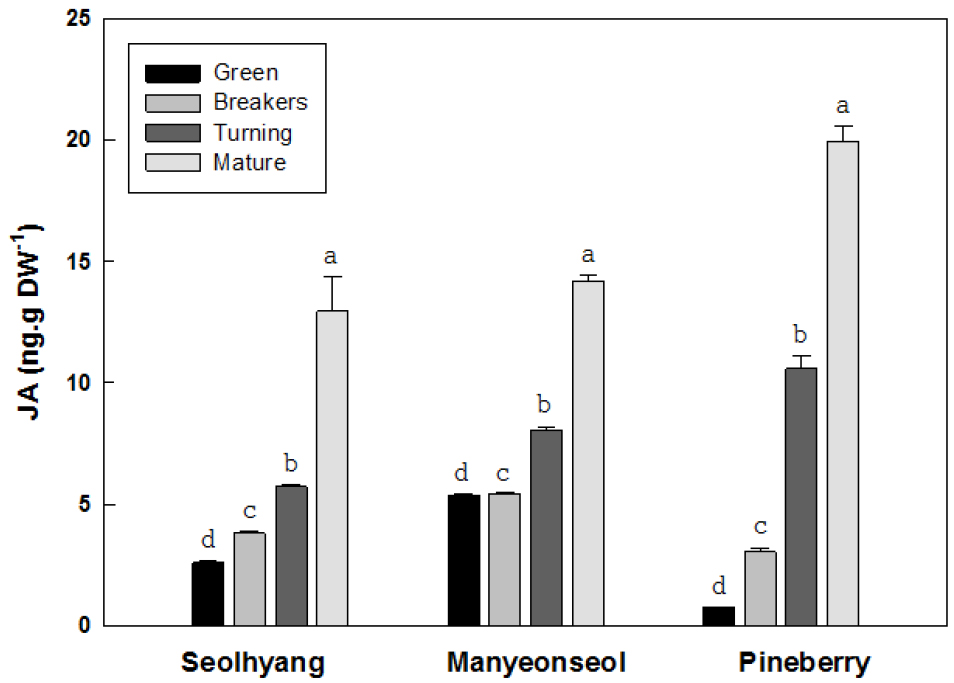
딸기 과실의 성숙단계별 자스몬산의 함량은 성숙과정이 진행될수록 증가하기 시작하여 완숙기에는 급격하게 증가하는 경향을 보였다(Fig. 7). 설향의 미숙기 자스몬산 함량은 건물 1g 당 2.59ng에서 변색기와 채색기에는 각각 3.82와 5.74ng으로 각각 1.5와 2.2배 증가하였고 완숙기에는 12.95ng으로 5배 증가하였다. 만년설의 미숙기 자스몬산 함량은 5.39ng에서 변색기와 채색기에는 각각 5.44와 8.05ng으로 증가하였으며 완숙기에는 14.19ng으로 2.6배 증가하였다. 파인베리의 미숙기 자스몬산 함량은 0.76ng으로 설향이나 만년설에 비해 아주 낮은 경향을 보였다. 그러나 변색기와 채색기의 함량은 각각 3.05와 10.58ng으로 4.0과 13.9배 증가하였고 완숙기의 함량은 19.94ng으로 미숙기 대비 26.2배 증가하였다.
7. 과실의 성숙단계에 따른 살리실산 함량 변화
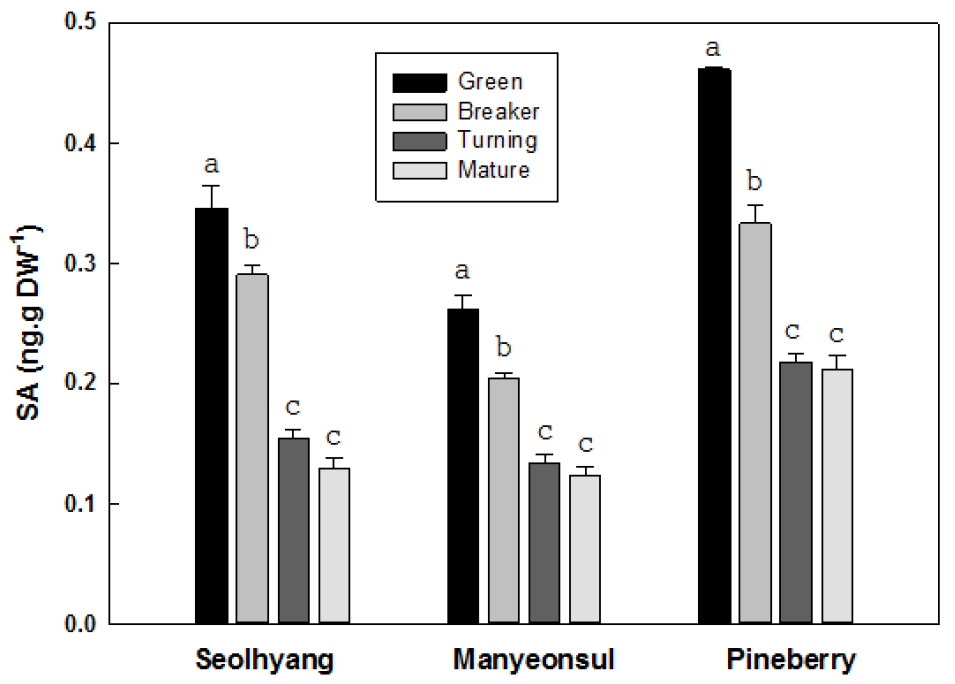
딸기 과실의 성숙단계에 따른 살리실산의 함량은 품종간에 차이는 있었지만, 전반적으로 모든 품종에서 미숙기의 함량이 가장 많았고 성숙이 진행될수록 감소하는 경향을 보였다(Fig. 8). 설향의 미숙기 살리실산 함량은 건물 1g 당 0.35ng에서 변색기와 채색기에는 각각 0.29과 0.16ng으로 15.9와 55.2% 감소하였고 완숙기에는 0.13ng으로 미숙기 대비 62.7% 감소하였다. 만년설의 미숙기 살리실산 함량은 0.26ng에서 변색기와 채색기에는 각각 0.20와 0.13ng으로 21.8과 48.7% 감소하였고 완숙기에는 0.12ng으로 52.9% 감소하였다. 파인베리의 미숙기 살리실산 함량은 0.46ng에서 변색기와 채색기에는 각각 0.33과 0.22ng으로 28.8과 52.9% 감소하였고 완숙기에는 0.21ng으로 미숙기 대비 54,2% 감소하였다.
고 찰
과실의 성숙과정에 생성되는 식물호르몬의 종류와 양은 다른데(Han 등, 2019; Li 등, 2019; Gupta 등, 2022), 일반적으로 성숙초기에는 발육과 생장에 긍정적인 영향을 미치는 호르몬이 생성되어 증가하지만, 성숙과정의 진행과 더불어 긍정적인 영향을 미치는 옥신, 시토키닌 및 지베렐린은 감소하는 반면에 노화를 촉진시키는 에틸렌과 앱시스산 함량은 증가한다(Leyser, 1998). 본 연구 결과에서도 딸기의 성숙 과정에서 옥신, 시토키닌, 지베렐린의 함량은 감소하였고, 앱시스산과 자스몬산은 증가하였다(Figs. 3, 4, 5, 6, 7). 옥신은 식물의 생장과 발육에 긍정적인 영향을 미치는 호르몬으로 세포분열과 더불어 과실의 발육에 중요한 역할을 하는데(Liu, 2019), 과실의 내생 옥신 함량은 착과기와 발육 초기에는 높지만 후기에는 감소한다고 하여 과실의 발육에는 영향을 미치지만 성숙에는 큰 역할을 하지 않는다(Zaharah 등, 2012; Teribia 등, 2016). 딸기 과실에서 옥신은 주로 종자에서 합성되어 화탁세포의 비대, 과실의 크기 및 과실의 성숙에 중요한 역할을 한다(Symons 등, 2012). 과실에서 종자를 제거하면 안토시아닌 함량이 감소한다는 보고도 있는데, 이는 옥신이 딸기 과실의 발육에 관여한다는 증거이기도 하다(Given 등, 1988). 설향, 만년설 및 파인베리의 성숙단계에 따른 IAA의 변화양상은 성숙과 더불어 증가한 후 채색기에서 가장 높았고 완숙기에는 감소하는 경향을 보였다(Fig. 3). 그러나 품종간의 함량 차이는 뚜렷하였는데, 설향과 만년설 과실의 채색기에는 건물 1g 당 각각 1,542와 1,487ng인데 비해 파인베리의 채색기에는 348ng으로 낮은 경향을 보였다. 딸기 3품종간 과실의 IAA 함량 차이는 과실의 크기와 비슷한 경향을 보였는데, 이러한 결과는 옥신이 과실의 생장과 비대에 크게 영향을 미친다는 보고와도 일치하였다(Given 등, 1988). 지베렐린은 줄기 생장, 종자의 발아, 개화 및 과실 생장을 촉진하고 과실의 단위결실을 유기하며 휴면을 타파하는 호르몬이다(Guleria 등, 2021). 딸기와 같은 호흡 비급등형 과실에서 지베렐린은 과실의 생장을 조절하는데(Symons 등, 2006), 과실의 발달과정에서 화탁세포의 비대는 내생 지베렐린에 의해서 조절된다(Csukasi 등, 2011; Martfnez 등, 1994). 딸기 과실에서 존재하는 지베레린은 주로 GA4와 GA1로 과실의 발달 초기에는 지베렐린 함량이 높지만, 성숙과 더불어 감소하는 것으로 알려져 있다(Csukasi 등, 2011; Symons 등, 2012). 특히 지베렐린은 옥신과 함께 과실의 착과와 발육에 관여하는 것으로 보고되었는데(Serrani 등, 2010; Bermejo 등, 2018; Hu 등, 2018; Shinozaki 등, 2020), 옥신은 세포분열에 관여하여 과실의 생장을 촉진하는 반면에 지베렐린은 IAA와 더불어 세포비대를 유도한다고 하였다(Liao 등, 2018). 본 실험에 사용한 딸기 품종간 성숙단계에 따른 지베렐린의 변화양상은 발육초기에 비해 성숙과 더불어 증가한 후 완숙기에는 감소하는 경향을 보여 Symons 등(2012)의 결과와 유사하였다. 그러나 성숙의 각 단계에 따른 품종간 차이는 과실의 크기 차이와 일치하여 과실이 컸던 설향과 만년설에서는 높은 반면 과실이 작았던 파인베리에서는 낮은 함량을 보였다(Fig. 4). 시토키닌은 옥신과 함께 세포주기를 조절하고 측아발달과 기관 형성에 영향을 미치며 노화를 지연시키는 호르몬이다(Guleria 등, 2021). 시토키닌은 옥신과 더불어 세포분열을 유도하여 과실의 생장을 촉진한다고 알려져 있다(Liao 등, 2018). 딸기에 있어서 시토키닌은 꽃눈분화에 관여하여 꽃수와 과실의 형성에 영향을 미치는데(Yamasaki와 Yamashita, 1993), 딸기 과실의 지아틴 농도는 개화기를 전후 하여 급격하게 증가하다가 발육이 진행될수록 감소한다(Eshghi와 Tafazoli, 2007). 본 실험에 사용한 딸기 과실의 성숙단계에 따른 지아틴 함량의 변화는 모든 품종에서 미숙기와 변색기에는 높다가 채색기 이후에는 급격하게 감소하여 Eshghi와 Tafazoli(2007)의 결과와 일치하였다(Fig. 5). 지아틴 농도가 가장 높았던 변색기에 있어서 설향, 만년설 및 파인베리의 지아틴 농도는 각각 건물 1g 당 3.2, 2.8 및 2.4μg에 비해 완숙기에는 각각 81, 82 및 83% 감소하는 경향을 보여 과실의 발육과 성숙에 대한 지아틴의 역할을 잘 반영해 주었다. 앱시스산은 옥신의 생장촉진과 지베렐린의 발아촉진 효과를 감소시키며, 노화와 휴면을 유기하는 호르몬으로 호흡 비급등형 과실의 성숙과정에 관여하는 중요한 호르몬이다(Symons 등, 2012; Karppinen 등, 2013; Guleria 등, 2021). 과실의 성숙은 과실 색, 구성성분, 경도 및 향기 등의 변화를 동반하는데 과실의 성숙과정에 일어나는 이러한 변화는 내생 옥신과 ABA에 의해서 일차적으로 시작된다(Perez-Llorca 등, 2019; Gupta 등, 2022). 옥신은 과실의 성숙과정을 지연시키는 반면에 ABA는 과실의 성숙과정을 촉진시키는 호르몬으로 옥신과 앱시스산의 상대적인 농도 차이에 의해서 성숙과정의 진행은 지연되기도 하고 촉진되기도 한다. 이러한 결과는 합성 호르몬 처리를 통하여 증명되었는데, 옥신계통의 합성호르몬인 NAA를 인위적으로 처리하면 딸기 과실에 안토시아닌 축적, 엽록소 소실 및 경도가 감소되어 성숙이 지연된다(Given 등, 1988). 그러나 앱시스산을 처리하면 과실의 연화와 더불어 적색 발현이 촉진되는데(Jiang과 Joyce, 2003), 외생 앱시스산은 PAL (phenylalanine ammonia lyase)의 활성을 유도하여 안토시아닌과 플라보노이드와 같은 페놀화합물의 생합성과 관련된 phenlpropanoid 대사과정을 활성화시킨다(Karppinen 등, 2018). 착색이 되지 않는 과실에서 앱시스산 함량이 감소하는데 이는 앱시스산 생합성에 중요한 역할을 하는 FaNCED1 유전자의 발현이 억제되기 때문이다(Jiang과 Joyce, 2003). Symons 등(2012)은 성숙하는 딸기 과실에서 앱시스산 함량이 증가하면 상대적으로 옥신과 지베렐린의 함량이 감소하여 연화나 착색과 같은 변화가 일어난다고 하였다. 또한 딸기 과실의 성숙과 더불어 종자에서 옥신의 합성이 감소하므로 과실의 성숙 과정이 조절(Given 등, 1988)된다고 하여 딸기 과실의 성숙과정에서 앱시스산과 옥신의 역할을 잘 반영하였다. 본 실험의 결과에서도 모든 품종에서 옥신 함량은 발육과 더불어 증가하다가 변색기 이후로는 급격하게 감소하는 경향을 보인 반면 앱시스산 함량은 성숙과정의 진행과 더불어 증가하는 경향을 보여 과실의 성숙과정에 관여하는 옥신과 앱시스산의 역할을 잘 반영하였다(Fig. 3, 6). 앱시스산 함량은 파인베리의 완숙기에 건물 1g 당 1.13µg인데 비해 설향과 만년설 과실에서 각각 4.13과 3.96µg으로 높은 경향을 보였다. 자스몬산의 생리적 기능은 스트레스에 대한 방어와 더불어 과실의 성숙과 노화를 촉진하는 것으로 알려져 있는데(Concha 등, 2013; Saavedra 등, 2016), 성숙이 진행되면 자스몬산의 농도는 증가하다가 완숙기가 되면 감소하는 것으로 알려져 있다(Kondo 등, 2000; Mukkun과 Singh, 2009). 딸기 미성숙과에는 자스몬산의 함량이 높지만 과실의 발육과 더불어 점차적으로 감소한다고 하였다(Gansser 등, 1997; Kondo 등, 2000; Kondo와 Fukuda, 2001). 미성숙한 딸기 과실에 자스몬산을 처리하면 호흡의 증가와 더불어 에틸렌이 생성되어 일시적으로 안토시아닌 색소가 합성되며 엽록소는 분해된다고 하여 자스몬산은 딸기 과실의 성숙에 관여한다고 하였다(Perez 등, 1997; Tao 등 2021). 또한 자스몬산을 처리하면 딸기와 토마토에서 안토시아닌과 리코펜 색소가 증가한다는 보고도 있는데(Perez 등, 1997; Liu 등, 2012), 이러한 색소는 딸기와 토마토에서 성숙의 지표로 활용된다는 점으로 보아 자스몬산은 과실의 성숙을 촉진시킨다고 할 수 있다. 본 실험에서는 3품종 모두 성숙과정이 진행될수록 자스몬산의 함량이 급격하게 증가하는 경향을 보여 미숙기 대비 완숙기에는 설향과 만년설에서는 각각 5배와 2.6배가 증가하였다(Fig. 7). 파인베리에서는 미숙기 대비 완숙기의 자스몬산 함량은 26.2배가 증가하여 건물 1g 당 19.94ng으로 가장 높은 경향을 보였다. 이러한 결과는 자스몬산 처리가 성숙을 촉진시킨다는 Liu 등(2012)의 결과와 일치하여 딸기 과실의 성숙에는 자스몬산이 중요한 역할을 하는 것으로 판단되었다. 특히 파인베리의 완숙과에서 나타나는 파인애플 향기는 자스몬산의 증가와 관련이 있을 것으로 보아 정밀한 분석을 통하여 기초자료를 수집한다면 차별화된 신품종 육성의 기초자료로 활용할 가치가 있을 것으로 판단되었다. 살리실산은 코리스믹산에서 유래된 식물호르몬으로 병원균에 의해서 과실의 수확 전후에 발생되는 피해를 방제하는 효과가 있다(Cao 등, 2013). 과실에 있어서 살리실산 함량은 과실의 발육단계에서는 높지만 성숙과 더불어 감소하는 것으로 알려져 있으며(Oikawa 등, 2015), 살리실산은 세포벽 분해효소의 활성을 억제하고 과피의 변색이나 호흡급등과 같은 성숙과정을 지연시킨다(Srivastava와 Dwivedi, 2000). 살리실산의 인위적 처리는 과실의 무게와 경도를 증가시키고 과색을 유지하는데 효과적이라는 보고도 있으며(Gimenez 등, 2014), 수확후 처리는 성숙을 지연시켜 품질을 유지한다(Zhang 등, 2003b; Valero 등, 2011). 살리실산은 ACC 산화효소의 발현을 억제함으로서 에틸렌과 상호작용하는데(Shi와 Zhang, 2012), 살리실산 처리에 의해서 ACC 산화효소의 활성이 증가하는 것은 에틸렌 합성은 살리실산에 의존적이며 아주 미세하게 조절된다고 하였다(Shi 등, 2013). 본 실험의 성숙단계에 따른 살리실산의 함량은 품종에 따른 함량 차이는 있었지만, 모든 품종에서 미숙기에서 가장 높았고 성숙이 진행될수록 감소하는 경향을 보였다(Fig. 8). 이러한 결과는 살리실산이 성숙과정을 지연시키거나(Srivastava와 Dwivedi, 2000) 과실의 경도를 증가시킨다는 보고(Zhang 등, 2003b; Gimenez 등, 2014) 등을 감안하면 과실의 성숙을 지연시키거나 경도가 높은 과실을 생산하기 위해서는 살리실산의 함량이 높은 품종을 개발하거나 인위적으로 처리하는 재배적인 방법도 검토할 필요가 있을 것으로 판단되었다.
