

서 론
온도구배하우스의 정의, 구조적 특징 및 제어 원리
1. 정의, 구조 및 미기상 환경
2. 열환경 형성 메커니즘 수리 모델
온도구배하우스의 기술적 발전 흐름
1. 1970년대: 자연광 기반 온도구배 실험 개념의 정립
2. 1980년대: 열수지 기반 온도구배 수리모델 정립을 위한 복사·열전달 특성 기반 확립
3. 1990년대: 온도구배 형성 원리의 체계화 및 환경 제어 기술의 발전
4. 2000년대 이후: 유입 공기 제어 기반 온도구배 시스템의 고도화와 검증
온도구배하우스 활용 기후변화 대응 작물 연구 사례
1. 온도에 따른 생육 및 발달 과정의 변화
2. 온도에 따른 생리적 스트레스 반응
3. 온도에 따른 수량 및 품질 반응
온도구배하우스의 구조적·기술적 한계와 주요 기후변화 연구시설 간 비교
온도구배하우스 이용 기후변화 대응 작물 연구 전망
서 론
기후변화에 관한 정부간 협의체(Intergovernmental Panel on Climate Change, IPCC)의 제6차 평가보고서에 따르면, 1850-1900년 대비 2011-2020년의 전 지구 평균 표면온도는 1.09°C 상승하였으며, 특히 1970년 이후의 온도 상승 속도는 지난 2,000년 동안 관측된 범위 중 가장 가파른 수준으로 보고되었다(IPCC 2023). 이러한 온난화는 평균 기온 상승에 그치지 않고 폭염, 가뭄, 집중호우 등 극한 기상 현상의 발생 빈도와 강도를 증폭시키며, 그 영향은 농업 생산 환경 전반에서 점차 뚜렷하게 나타나고 있다.
기후변화 조건에서 작물은 생육 기간 동안 반복적인 고온 스트레스와 수분 불균형에 노출되며, 이에 따라 생육 반응과 수량 형성이 불균등하게 나타나 생산성의 변동성과 불확실성이 증가하는 것으로 보고되고 있다(Rosenzweig et al. 2014). 특히 강수 패턴의 불균등성 심화는 가뭄과 홍수 발생 빈도를 증가시켜 작물 수확량 감소로 직결된다(Yuan et al. 2024). 이러한 기후 변동성과 극한 기상 현상은 농업 생산 시스템 전반에서 상당한 경제적 손실을 초래하는 것으로 분석되고 있다(Habib-ur-Rahman et al. 2022).
기후변화의 영향은 모든 작물과 재배 지역에서 동일하게 나타나지 않으며, 작물의 생리적 특성, 생육 단계, 재배 환경에 따라 반응 민감도가 다르게 나타난다. 또한 온난화에 따른 생물계절 변화는 개화 및 생육 시기를 앞당기고, 작기 조정과 재배 적지의 공간적 이동을 유도함으로써 농업 생산 체계 전반의 구조적 변화를 초래한다(Wolfe et al. 2005; RDA 2016; Kim and Kim 2018).
이와 같은 변화는 단순한 수확량 변동을 넘어, 환경 요인이 작물 생육 과정의 어떠한 경로와 생육 단계에서 생산성에 영향을 미치는지를 규명할 필요성을 제기한다. 특히 기후변화 조건에서는 온도, CO2, 수분과 같은 개별 환경 요인이 생육 기간 전반에 걸쳐 누적되거나 특정 생육 단계에 집중적으로 작용함에 따라, 작물 반응이 단계별로 상이하게 나타난다(Vu and Kim 2011; Liu et al. 2025). 따라서 기후변화가 작물 생육과 생산성에 미치는 영향을 정량적으로 분석하기 위해서는 개별 환경 요인의 영향을 분리하여 평가할 수 있는 실험적 접근이 요구되어 왔다.
이러한 연구 목적에 대응하여 다양한 환경조절 실험시설이 단계적으로 개발·활용되어 왔다. 생장상(Growth Chamber)은 온도와 광 환경을 정밀하게 제어할 수 있어 작물의 기초 생리 반응을 규명하는데 활용되어 왔으며, 이후 출입형 생장상(Walk-in Growth Chamber)은 재배 공간을 확장함으로써 생장상의 공간적 제약을 일부 완화하고자 개발되었다. 나아가 토양-식물-대기 연구 시스템(Soil-Plant-Atmosphere-Research system, SPAR)은 자연광 조건을 유지한 상태에서 환경 제어가 가능하도록 설계되어 생장상 기반 실험의 현실성을 보완하였다(Reddy et al. 2001). 그러나 실험시설들은 여전히 공간 규모가 제한적이어서 작물 군락 구조와 포장 조건을 충분히 재현하는 데에는 구조적 제약이 존재하는 것으로 보고되고 있다(Poorter et al. 2016).
한편 개방형 상부 챔버(Open-Top Chamber, OTC)와 자유대기 CO2 농도 증강 시설(Free-Air CO2 Enrichment, FACE)은 자연광과 야외 조건을 유지할 수 있다는 장점이 있다. 그러나 OTC의 구조적 제약과 FACE의 바람 의존적 CO2 농도 조절 특성으로 인해 온도 상승의 정밀 제어와 CO2 및 온도 효과의 독립적 분리 평가에는 기술적·경제적 한계가 존재하는 것으로 보고되어 왔다(Seo et al. 2016). 이와 함께 생태 전이지대 실험시설(Ecotone), 생물군계 실험시설(Biome), 지구변화 실험시설(Global Change Experimental Facility) 등 대형·복합 환경조절 시설들이 구축되어 생태계 수준의 기후변화 영향 분석에 활용되어 왔다. 그러나 이러한 시설의 경우도 동일한 광환경에서 작물의 연속적인 온도 반응을 체계적으로 비교·분석하는 데에는 적용상의 제약이 있는 것으로 인식되어 왔다(Schädler et al. 2019; Lee et al. 2022).
온도구배하우스(Temperature Gradient Greenhouse, TGG)는 온도 요인에 초점을 두고 작물의 생육 및 생리 반응을 정량적으로 분석하기 위해 개발된 기후변화 연구시설이다. TGG는 단일 시설 내에서 길이 방향을 따라 연속적인 온도 구배를 형성함으로써, 동일한 자연광 조건을 유지한 상태에서 상이한 기온 수준에 따른 작물 반응을 동시에 비교·분석할 수 있는 구조적 특성을 지닌다(Morales et al. 2014). 이와 같은 구조적 특성은 동일한 광환경 하에서 온도 효과를 상대적으로 분리하여 평가할 수 있게 하며, 작물의 온도 반응 곡선과 임계 반응 구간을 정량적으로 도출하는 데 적합한 실험 환경을 제공한다(Allen et al. 2020).
본 논문은 기후변화 대응 작물 연구를 위한 실험시설 중 하나인 TGG에 주목하여, 그 구조적 원리와 열환경 형성 메커니즘, 기술적 발전 과정, 그리고 국내외 연구 사례를 종합적으로 고찰하고자 한다. 아울러 TGG의 구조적·기술적 한계를 분석하고 SPAR, FACE, OTC 등 주요 기후변화 연구시설과의 비교를 통해 TGG의 상대적 특성과 적용 범위를 정리하며, 향후 작물 모델 검증 및 복합 환경 실험을 위한 연구 인프라로서의 활용 전망을 논의하고자 한다.
온도구배하우스의 정의, 구조적 특징 및 제어 원리
1. 정의, 구조 및 미기상 환경
TGG는 자연광 조건에서 단일 온실 내부에 연속적인 공기 온도 구배를 형성하도록 설계된 기후변화 연구용 실험시설이다. 이를 통해 동일한 광환경에서 상이한 기온 조건에 대한 작물의 생육 및 생리 반응을 동시에 비교·분석할 수 있다 (Hadley et al. 1995; Okada et al. 1995; Morales et al. 2014; Lee et al. 2022). 이러한 실험 개념은 1970년대 이후 기후변화에 따른 작물 반응을 정량적으로 규명하기 위한 목적에서 제안되었으며, 이후 국내외에서 다양한 형태로 적용·발전되어 왔다(Fig. 1).

Fig. 1
Representative temperature gradient facilities utilized for climate change-related crop studies: (A) TGG at the National Agriculture and Food Research Organization (NARO), Japan, (B) TGG at the Rural Development Administration (RDA), Korea, and (C) TGG operated by the Jeonbuk State Agricultural Research & Extension Services, Korea. Images in (A) and (B) were reproduced from the respective institutional websites (NARO, https://www.naro.go.jp/laboratory; RDA, https://www. rda.go.kr)
TGG는 일반적으로 상용 비닐하우스 구조를 기반으로 한 터널형 온실로 구축되며, 선행 연구들에서는 폭 약 3-4m, 길이 약 20-30m의 장축형 구조가 주로 사용되어 왔다(Hadley et al. 1995; Okada et al. 1995). 하우스의 일측 끝단에는 외기 유입부가, 반대편 끝단에는 배기팬을 포함한 유출부가 설치되어 내부 공기 흐름을 장축 방향으로 유도한다. 유입된 외기는 하우스 내부를 이동하면서 태양 복사 에너지와 보조 가열에 의한 현열을 흡수하며 점진적으로 가열되고, 그 결과 유입부에서 유출부로 갈수록 기온이 단계적으로 상승하는 연속적 온도 구배가 형성된다(Okada et al. 1995). 국내에서도 농촌진흥청 국립원예특작과학원 등 주요 연구기관을 중심으로 이러한 설계 원리를 기반으로 한 TGG가 구축·운영되어 왔다.
전북특별자치도농업기술원에 구축된 TGG는 길이 35m, 폭 6.5m, 높이 3.5m의 단동 비닐하우스 구조이며, 유입부와 유출부의 배치 및 환기 장치 구성은 위의 기본 설계 개념을 따르고 있다(Fig. 2). 해당 시설에서는 유입부로부터 장축 방향을 따라 각각 T1(7m), T2(19m), T3(31m) 지점에 온도 센서를 설치하여 내부 미기상 환경을 관측하였다. 그 결과, 유입부 인근(T1)에서 유출부 인근(T3)으로 갈수록 공기 온도가 단계적으로 증가하는 공간적 분포가 일관되게 확인되었다(Fig. 3). 이러한 미기상 특성은 TGG에서 공통적으로 보고되는 핵심적인 환경적 특징에 해당한다(Lee et al. 2022).
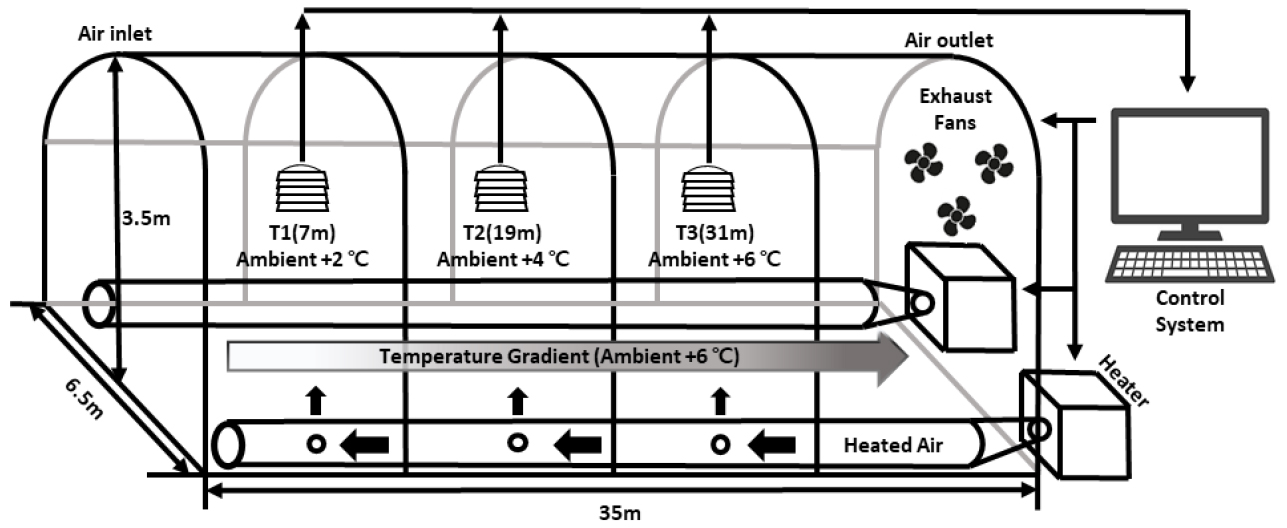
Fig. 2
Schematic diagram of the TGG. The facility consists of a single-span plastic greenhouse (35m length × 6.5m width × 3.5m height) in which a unidirectional airflow is established along the longitudinal axis. Ambient air enters through the inlet at one end of the greenhouse and is continuously exhausted by ventilation fans installed at the opposite end, forming a continuous air temperature gradient of up to +6 °C under natural solar radiation
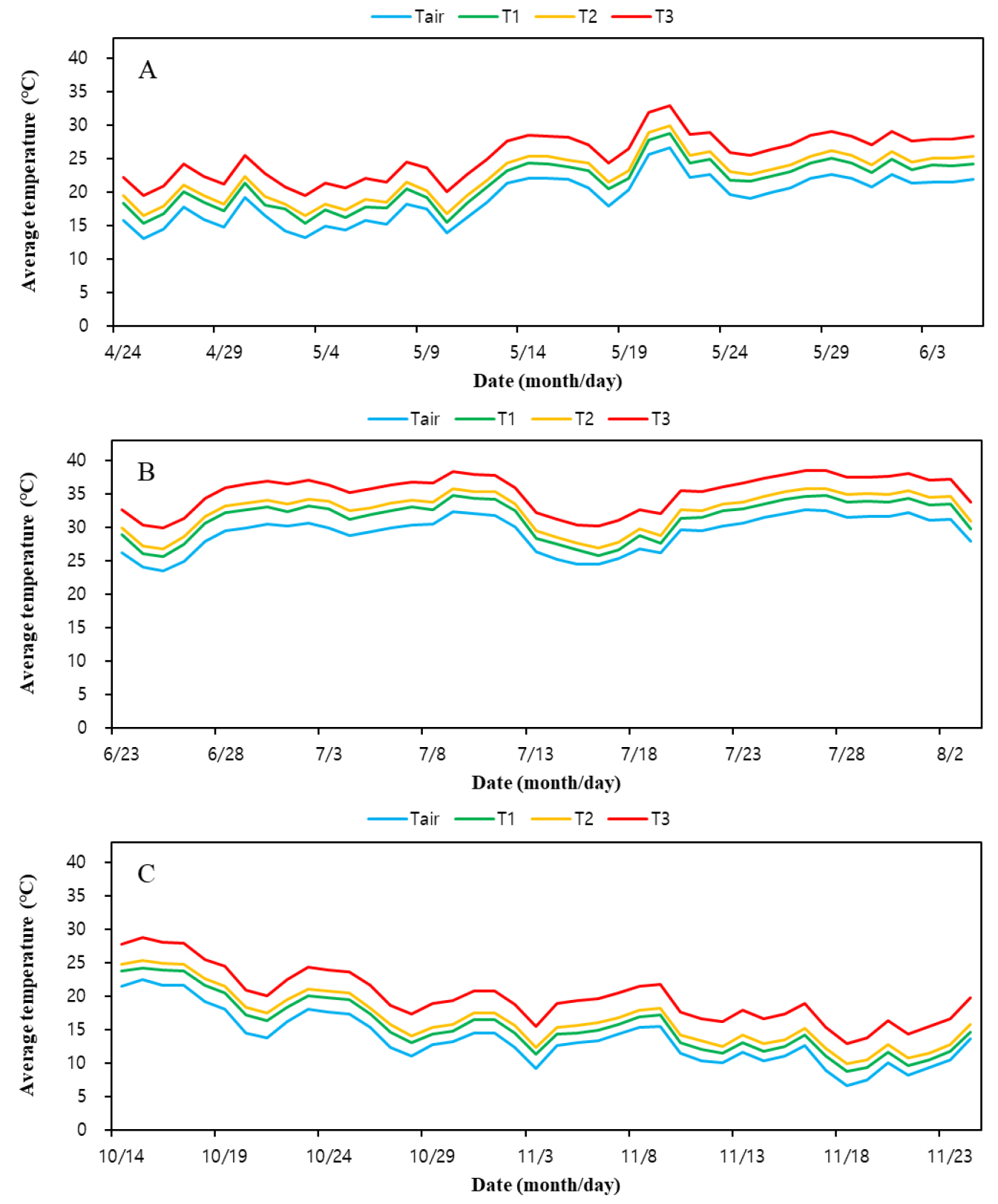
Fig. 3
Spatial distribution of air temperature within the temperature gradient greenhouse under natural light conditions. Air temperature was measured at the inlet (Tair) and at three positions along the longitudinal axis (T1-T3). Panels (A), (B), and (C) represent spring, summer, and autumn, respectively
2. 열환경 형성 메커니즘 수리 모델
TGG 내부의 온도 구배(ΔT) 형성은 태양 복사 에너지 유입에 의한 현열 획득(H)과 강제 환기에 의한 공기 수송(V) 사이의 에너지 수지(Energy balance) 기작에 의해 결정된다. 하우스 유입구로 도입된 외기는 지면과 작물체로부터 발생하는 현열을 흡수하며 배출구 방향으로 이동하고, 이 과정에서 공기 흐름의 경로를 따라 점진적인 기온 상승이 유도된다. 이러한 열역학적 관계는 Okada et al.(1995)에 의해 다음과 같은 수리 모델로 공식화되었다.
여기서 ΔT는 TGG 공기 유입구와 유출구 사이의 공기 온도 차이(℃)이며, H는 단위 지면적 기준의 현열 획득량(W·m-2)이다. Cp는공기의 비열(1004J·kg-1·℃-1), ρ는 공기 밀도(1.2 kg·m-3), V는 단위 지면적당 환기율(m3·m-2·s-1)이다. 또한 α는 피복 면적(지붕 및 벽면)의 지면적 대비 비율이며, ht는 피복재를 통한 열전달계수(W·m-2·℃-1)이다. 계수 0.5는 시설 내부 평균 공기 온도를 유입구와 유출구 공기 온도의 산술평균으로 가정함에 따라 도입되었다(Okada et al. 1995).
이 수리모델에 따르면 환기율이 감소할수록 온도 구배는 증가하나, 환기량이 불충분할 경우에는 강제 환기에 의한 단방향 기류보다 자연대류의 영향이 우세해질 수 있다. 이는 TGG 내 온도 구배의 폭과 공간적 분포가 환기 효율 및 열수지 간의 물리적 균형에 의해 결정됨을 의미한다. 따라서 해당 모델은 TGG의 설계 및 제어 전략을 이해하기 위한 이론적 틀을 제공하지만, 실제 구배 크기와 공간적 분포는 외기 조건, 피복재 특성, 작물 군락 구조에 따라 달라질 수 있어 현장 검증을 통해 보완적으로 해석될 필요가 있다.
온도구배하우스의 기술적 발전 흐름
TGG는 자연광 조건에서 온도의 단독 효과를 규명하기 위한 실험 개념으로 출발하여, 연구 목적의 확장과 실험적 한계에 대한 기술적 보완을 거치며 단계적으로 발전해 왔다(Table 1). 초기에는 온도구배 형성의 물리적 가능성을 탐색하는 데 초점이 맞추어졌으나, 이후 열환경 형성 원리에 대한 이론적 정립과 환경 제어 기술의 도입을 통해 활용 범위가 확장되었으며, 이 과정에서 시설의 구조적 설계 또한 진전되었다(Fig. 4).
Table 1
Technological development of temperature gradient greenhouse systems
| Period | Technological focus | Technical outcomes | Key references |
| 1970s | Establishment of the concept of natural-light-based temperature gradient experiments | Introduction of continuous temperature-gradient formation under natural light, enabling separation of temperature effects from other environmental factors | Mihara(1971) |
| 1980s | Establishment of radiative and heat-transfer properties for heat-balance-based temperature-gradient modeling | Quantification of radiative properties of covering materials, providing a physical basis for heat-balance analysis and modeling | Okada(1983) |
| 1990s | Systematization of temperature-gradient formation principles, development of environmental control technologies, and platform-level validation | Establishment of heat-balance-based temperature-gradient formation principles, development of control strategies addressing spatial non-uniformity in large facilities, and experimental validation of combined temperature-CO2 manipulation under natural light | Okada et al.(1995); Sinclair et al.(1995); Hadley et al.(1995); Horie et al.(1995) |
| 2000s-present | Advanced temperature-gradient systems based on inlet-air control, multi-factor integration, and quantitative validation | Independent control of inlet air temperature, implementation of combined temperature-CO2 gradients, integration of water availability control for multi-factor experimentation, advancement of precision control, and transition to CFD-based quantitative validation | Okada et al.(2000); Lee et al.(2001); Lee(2011); Morales et al.(2014); Seo et al.(2021) |
1. 1970년대: 자연광 기반 온도구배 실험 개념의 정립
TGG 시스템의 학술적 기원은 Mihara(1971)가 제안한 온도 반응 곡선(Temperature Response Curve) 실험 개념에서 비롯된다. 초기 형태는 높이 50cm, 길이 10m의 소형 플라스틱 튜브 온실 구조로, 별도의 인공 공조 장치 없이 태양 복사 에너지 유입과 환기 속도 사이의 에너지 평형을 통해 하우스 길이 방향의 온도 구배를 형성하였다. 이러한 접근은 자연광 조건에서 외기 기온의 일변화에 따른 온도의 단독 효과를 분리·관찰할 수 있는 실험적 틀을 제시하였으며, 이후 TGG가 기후변화 대응 작물 연구 시설로 확장되는 개념적·기술적 출발점을 마련하였다.
2. 1980년대: 열수지 기반 온도구배 수리모델 정립을 위한 복사·열전달 특성 기반 확립
TGG의 수리 모델을 정교화하는 과정에서 피복재의 복사 특성을 규명한 연구는 중요한 이론적 전환점을 제공하였다. Okada(1983)는 방사계법(Emissiometer method)을 이용하여 온실 피복용 플라스틱 필름의 장파복사 특성을 정량적으로 측정하였으며, 이를 통해 열수지 계산에 필요한 복사 매개변수를 실측 기반으로 제시하였다. 이러한 물성 정보의 확립은 이후 TGG 내부 열환경을 에너지 수지 관점에서 해석하고 수리적으로 모델링하는 데 핵심적인 기반을 제공하였다.
3. 1990년대: 온도구배 형성 원리의 체계화 및 환경 제어 기술의 발전
TGG의 내부의 열환경 형성 메커니즘이 수리적으로 정립되는 한편, 시설 대형화에 따른 공간 불균일성과 외기 변동성에 대응하기 위한 환경 제어 기술이 본격적으로 도입되었다. Okada et al.(1995)는 TGG를 대상으로 환기율, 복사 조건, 현열 획득 간의 상호작용을 열수지 관점에서 체계적으로 제시함으로써, 안정적인 공기 온도 구배가 형성되는 물리적 기반을 확립하였다(Fig. 4A). 이러한 이론적 기반 위에서 Sinclair et al.(1995)는 시설 내부를 구획화하고 패들 팬을 설치하여 수직·수평 혼합을 강화함으로써, 플롯 단위의 독립적인 온도 환경 유지가 가능함을 보고하였다. 또한, Hadley et al.(1995)는 이중벽 구조와 분리형 히트펌프 기반의 능동적 냉·난방 시스템을 적용하여, 외기 변동과 무관하게 안정적인 온도구배 유지를 가능하게 하였다.
이러한 기술적 진전을 바탕으로 Horie et al.(1995)는 TGG에서 온도 조절과 CO2 농도 상승 처리를 결합하여, 자연광 조건에서 온도와 CO2를 동시에 처리할 수 있는 기후변화 실험이 기술적으로 가능함을 실증하였다(Fig. 4B).
4. 2000년대 이후: 유입 공기 제어 기반 온도구배 시스템의 고도화와 검증
TGG 시스템은 유입 공기 온도를 외기 조건과 분리하여 제어하는 방식으로 발전하였다. Okada et al.(2000)는 사전 공조를 도입한 TGG를 제시하여, 고온기에도 안정적인 온도구배 형성이 가능함을 보였다. 아울러 천공 덕트 기반 공기 분배 구조를 통해 기류 역류 문제를 구조적으로 완화하였다(Fig. 4C). 이러한 유입 공기 제어 기술을 바탕으로 Lee et al.(2001)는 시설 길이 방향으로 온도와 CO2 농도를 동시에 선형적으로 변화시킬 수 있는 환경을 구현하여, 복합 기후 요인의 상호작용을 단일 실험 환경에서 분석할 수 있는 연구 시설을 제시하였다.
이후 Morales et al.(2014)은 기존의 온도와 CO2 제어 기반 온도구배 시스템에 수분 가용성 제어를 추가하여, 다요인 기후변화 연구 시설로 확장하였다. 이들은 TGG 내에서 CO2 농도 조절, 온도구배 형성, 토양 센서 기반 관수 제어를 결합함으로써 기후 요인 간 상호작용을 분석할 수 있도록 하였다. 또한 생장 챔버로 전환 가능한 개념을 도입하고, TGG 내 화분을 토양 내에 배치하여 근권부 온도 변동이 보다 자연 조건에 가깝게 나타나도록 하였다. 국내에서는 이후 TGG의 제어 정밀도를 더욱 향상시키는 기술적 진전이 이루어졌다. Lee(2011)가 등록한 특허 기술은 유입구 메시 구조를 통한 기류 균일화와 비례·적분·미분(PID) 제어 기반의 보조 난방 시스템을 결합함으로써, 일변화 조건에서도 주·야간 연속적인 온도구배 유지를 가능하게 하였다.
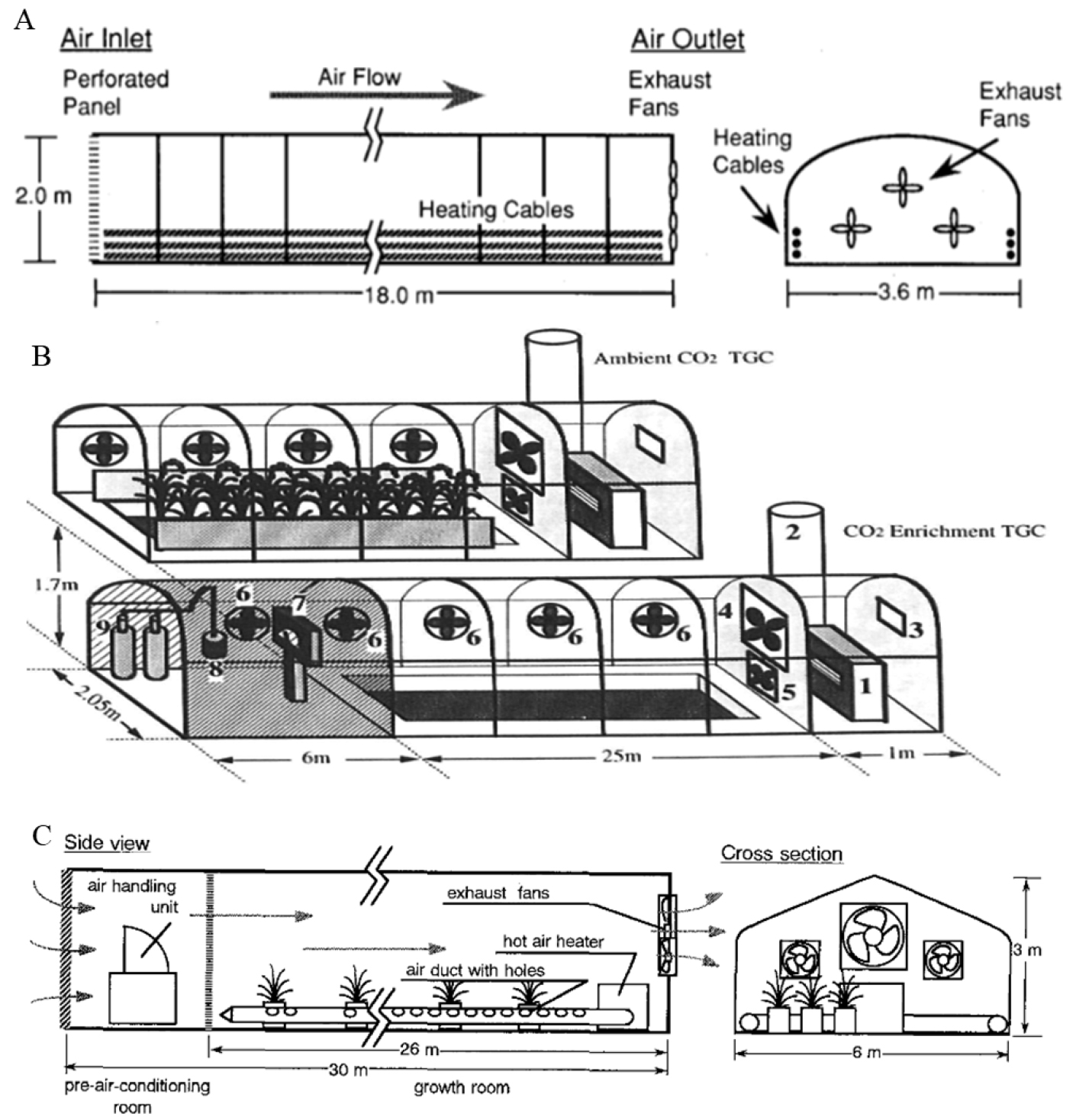
Fig. 4
Representative designs illustrating the technological evolution of temperature gradient greenhouses: (A) a basic temperature gradient greenhouse based on inlet-outlet ventilation, (B) a temperature-CO2 combined temperature gradient greenhouse under natural light conditions, and (C) a pre-air-conditioned temperature gradient greenhouse with controlled inlet air temperature. Figures in (A), (B), and (C) were reproduced from Okada et al.(1995), Horie et al.(1995), and Okada et al.(2000), respectively
최근에는 전산유체역학(Computational Fluid Dynamics, CFD)과 다지점 온도 센서를 결합한 공간 해석을 통해 TGG 내부의 기류 분포와 열환경 구조를 정량적으로 검증하려는 연구가 이루어지고 있다. Seo et al.(2021)은 90개 지점의 실측 온도 자료를 이용하여 CFD 결과를 검증하였으며, 0.8m 높이에서 평균오차 0.597°C, 길이방향 평균 RMSE 0.103°C를 제시하였다. 이러한 결과는 TGG 내 열환경에 대한 수치 모의의 신뢰성을 뒷받침하며(Fig. 5), CFD 기반 공간 해석이 설계 원리와 제어 전략의 물리적 타당성을 평가하고 온도구배 실험의 재현성을 높이는 데 활용될 수 있음을 보여준다.
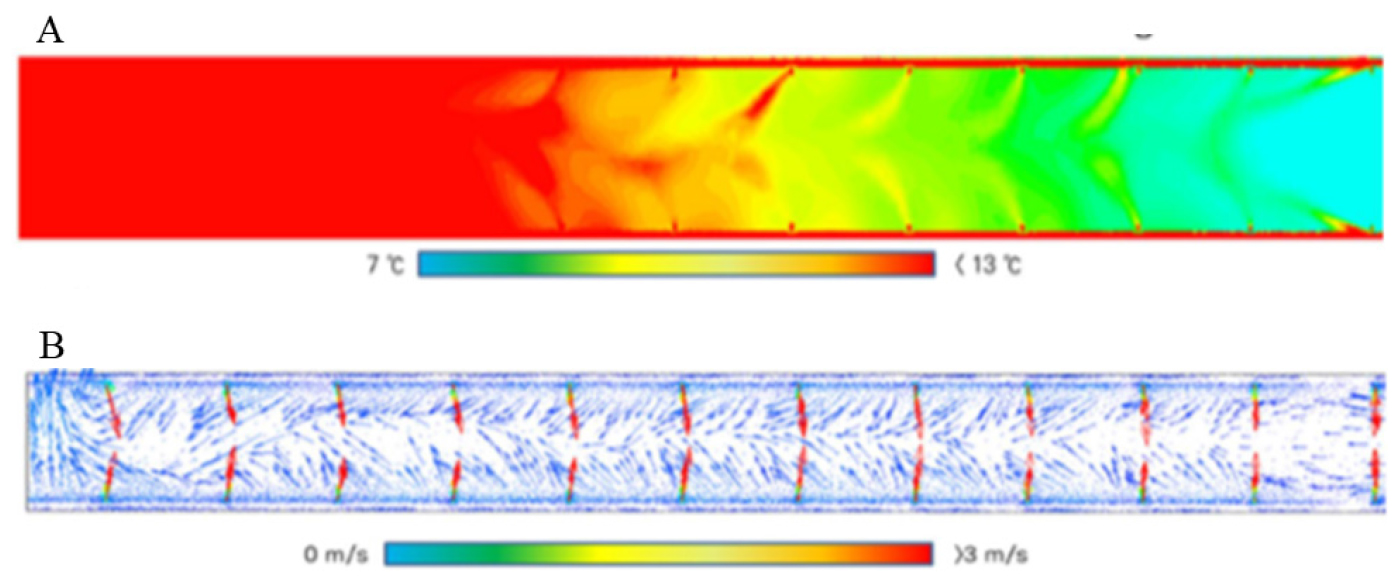
Fig. 5
CFD-simulated air temperature distribution (A) and airflow patterns (B) in a temperature gradient greenhouse. Reproduced from Seo et al. (2021)
온도구배하우스 활용 기후변화 대응 작물 연구 사례
TGG를 활용한 연구들은 온도 상승이 작물의 생육 및 발달 과정, 생리적 스트레스 반응, 그리고 최종 수량과 품질 형성에 이르기까지 연속적이고 단계적인 영향을 미침을 보고하였다. 온도 변화는 먼저 생육 속도와 발달 단계의 진행을 조절하며, 임계 온도 초과 시 생리적 스트레스 반응을 유발하고 광합성 기능을 저해하였다. 이러한 생리적 반응은 동화산물 분배 효율과 저장 기관 기능 저하로 이어져 수량 및 품질 감소로 연결되었다(Table 2).
Table 2
Major research themes and key findings from crop studies using temperature gradient-based facilities
| Research focus | Key findings | Representative crops | Key references |
|
Growth and developmental responses to temperature | Rising temperature accelerated growth and developmental progression (e.g., leaf appearance, flowering), while supra-optimal temperatures caused developmental delay or abnormal growth depending on crop and ecotype |
Alfalfa Chickpea, Garlic, Rice, Wheat, Chinese cabbage, Soybean | Aranjuelo et al.(2005); Chakrabarti et al.(2013, 2021); Kim et al.(2023); Oh et al.(2019); Oh et al.(2023); Rawson et al.(1995); Seo et al.(2021) Tacarindua et al.(2012, 2013); |
|
Physiological stress responses under high temperature | High temperature reduced photosynthetic capacity and stomatal conductance, increased water and oxidative stress, and showed clear differences among crops and ecotypes |
Grapevine, Tomato, Potato, Soybean, Rice, Black gram, Chinese cabbage | Kizildeniz et al.(2018); Laxman et al.(2018); Lee et al.(2024); Liu et al.(2025); Oh et al.(2023); Pasham et al.(2025) Seo et al.(2021); |
|
Yield and quality responses to temperature stress | Heat stress constrained assimilate partitioning and sink development, leading to reduced yield components and quality deterioration, particularly under reproductive-stage exposure |
Soybean, Potato, Rice, Black gram Chinese Cabbage, Adzuki bean | Baek et al.(2020); Tacarindua et al.(2012, 2013); Lee et al.(2024); Oh et al.(2023); Pasham et al.(2025); Seo et al.(2021); Suh et al.(2023); |
1. 온도에 따른 생육 및 발달 과정의 변화
식물의 발달 속도는 온도 변화에 대한 생리적 반응의 민감도에 의해 결정되며, 일반적으로 생육도일 및 광열단위의 누적량에 비례하여 가속화되는 경향을 보인다(Chakrabarti et al. 2021). 생육도일의 축적은 작물의 마디 생성 및 엽 출현 속도를 결정하는 주요 요인이며, 결과적으로 개화 및 성숙과 같은 생육 단계에 도달하는 시기를 단축시킨다(Rawson et al. 1995). TGG에서 알팔파, 밀, 병아리콩, 마늘, 벼, 배추를 대상으로 수행된 연구들에 따르면, 온도 상승에 따라 생육 및 발달 과정이 촉진되었다(Aranjuelo et al. 2005; Chakrabarti et al. 2013; Oh et al. 2019; Seo et al. 2021; Oh et al. 2023).
특히 배추의 경우 외기온도 보다 약 3°C 높은 온도 조건에서 엽수는 약 26.8%, 엽면적은 약 45.8% 증가하며 왕성한 생육 반응을 보였다(Seo et al. 2021). 그러나 특정 임계 온도를 초과하는 고온 환경에서는 생육 저해 현상이 관찰되었다. 콩의 경우 외기 대비 +3°C 이상의 온도구간에서 개화기부터 종자 형성기까지의 기간이 5일 이상 지연되는 발달 정체 현상이 확인되었다(Tacarindua et al. 2012, 2013). 또한 마늘에서 생태형에 따라 상이한 온도 반응을 나타냈다. 난지형의 경우 외기 대비 3°C 이상의 온도구간에서 추대 기간이 10일 단축 되어 발달이 촉진되었으나(Oh et al. 2019), 한지형 마늘에서는 외기 대비 3°C 이상의 온도 구간에서 불완전 추대율이 약 25.6%까지 급증하여 정상적인 발달 과정이 저해되는 결과는 나타냈다(Kim et al. 2023). 이러한 결과는 온도에 따른 생육 및 발달 반응이 단순한 선형적 촉진이 아니라, 작물 및 생태형에 따라 임계점을 갖는 비선형적 특성을 보여준다.
2. 온도에 따른 생리적 스트레스 반응
온도 상승은 식물의 세포막 유동성을 변화시키고 광계 II의 광화학적 효율을 감소시켜 생리적 스트레스를 유발한다(Sunoj et al. 2016; Sun et al. 2018). 특히 고온에 의한 기공전도도의 불균형은 증산 조절 이상과 수분이용효율의 저하를 야기하며, 이는 활성산소 축적과 지질 과산화 증가에 따른 대사작용 교란으로 이어진다(Yuan et al. 2017; Rangappa et al. 2024). 이러한 생리적 장애는 광합성 능력을 저하시켜 탄소 동화 과정에 심각한 저해를 유발한다.
TGG를 이용한 연구에서 토마토와 벼는 고온 노출 시 순광합성률과 기공전도도가 감소하여 탄소 동화 능력이 감소되는 경향을 보였으며(Laxman et al. 2018; Oh et al. 2023), 포도, 감자, 콩, 녹두, 배추에서도 고온 스트레스에 따른 기공 조절 기능 약화와 수분 스트레스 증가가 공통적으로 관찰되었다(Kizildeniz et al. 2018; Seo et al. 2021; Lee et al. 2024; Liu et al. 2025; Pasham et al. 2025). 이러한 수분스트레스 조건에서는 광합성 기구의 구조적·기능적 손상으로 이어져 탄소 동화 효율 저하를 유발하는 주요 요인으로 작용하였다. 특히 감자의 경우, 저온 적응형 생태형인 ‘수미’는 외기 대비 1.6°C 이상의 온도 상승 구간에서 엽록소 함량과 광합성 효율이 감소하고, 전 온도 구간에서 높은 작물 수분 스트레스 지수를 나타내 고온에 대한 생리적 적응 한계를 나타냈다. 반면 ‘조풍’은 외기 대비 3.5°C 수준까지의 온도 상승 조건에서도 광합성 효율을 안정적으로 유지하여 상대적으로 높은 내열성을 나타냈다(Lee et al. 2024). 이러한 결과는 고온에 대한 생리적 스트레스 반응이 작물뿐만 아니라 생태형 수준에서도 차별적으로 나타남을 확인하였다.
3. 온도에 따른 수량 및 품질 반응
작물의 최종 생산성은 광합성 산물이 저장 기관으로 전이되는 분배 효율에 의해 결정된다(Tacarindua et al. 2012). 고온 스트레스는 동화 산물의 전류 및 분배 과정을 저해할 뿐만 아니라, 저장 기관의 형성 및 수용 능력 자체를 생리적으로 제한하여 생장속도와 성숙 기간 사이의 불일치를 초래한다(Baek et al. 2020).
TGG에서 녹두, 팥, 콩은 고온 노출 시 꼬투리 수와 종자 수가 급격히 감소하며 수확 지수가 저하되는 양상을 보였고(Tacarindua et al. 2012, 2013; Suh et al. 2023; Pasham et al. 2025), 배추와 감자에서는 적정 온도를 초과할 경우 결구 및 괴경 비대가 억제되어 상품성이 크게 훼손되는 결과가 공통적으로 관찰되었다(Seo et al. 2021; Lee et al. 2024).
특히 벼의 경우 개화기 고온 노출로 인한 화분 임성 저하가 이삭 불임률 증가로 이어졌으며, 이는 동화 산물이 결실 기관으로 전이되지 못하고 영양기관에 잔존하는 현상을 유발하였다(Oh et al. 2023). 그 결과 출수 후기까지 엽록소 함량과 식생지수가 높게 유지되는 지연된 노화 현상이 관찰되어, 고온 조건이 수량 감소와 생리적 노화 양상의 변화에 동시에 영향을 미치는 것으로 확인되었다. TGG에서 온도 상승은 작물의 수량 및 품질에 대해 임계 온도를 갖는 비선형적 반응을 나타내며, 임계 온도 초과 시 동화산물 분배 저해와 저장 기관 발달 제한이 동시에 발생하여 수량 감소와 품질 저하로 이어졌다.
온도구배하우스의 구조적·기술적 한계와 주요 기후변화 연구시설 간 비교
TGG는 장축 방향으로 약 4-6°C 범위의 연속적 온도 구배를 안정적으로 형성할 수 있는 수준까지 발전해 왔다(Sinclair et al. 1995; Okada et al. 2000). 그러나 TGG 내부에서는 가열된 공기의 부력 효과와 강제 환기 패턴의 상호작용으로 수직 방향의 온도 불균일성이 발생한다. Seo et al.(2021)은 높이 0.4, 0.8, 1.2m에서의 온도차가 각각 4.60, 4.52, 5.62°C로 나타남을 확인하였고, Horie et al.(1995)은 수직 성층화를 완화하기 위해 측벽에 진동팬을 설치하였다. 아울러 온도 구배는 포화수증기압의 공간적 차이를 유발하여 수증기압포차 구배를 수반한다 수반하며, Morales et al.(2014)은 +4°C 처리구에서 야간 상대습도가 유의하게 감소함을 보고하였다. 따라서 TGG에서 관측되는 작물 반응은 순수한 온도 효과뿐 아니라 습도 변화가 결합된 반응일 가능성을 함께 고려하여 해석할 필요가 있다(Okada et al. 1995).
또한 TGG는 생장상보다 자연광을 이용한다는 관점에서 포장 조건에 가깝지만, 피복재와 시설 구조에 따른 미기상 차이가 존재한다. Morales et al.(2014)은 시설 내부의 광합성유효복사가 외부 대비 약 30-40% 감소한다고 보고하였으며, Horie et al.(1995)은 내부 풍속이 계절 평균 0.22m·s⁻¹ 수준으로 포장보다 현저히 낮다고 제시하였다. Poorter et al.(2016) 역시 제어환경과 포장 사이의 광, 스펙트럼, 기류 등의 구조적 차이가 식물 반응에 영향을 줄 수 있음을 언급한 바 있다. 따라서 TGG는 포장 유사 실험환경으로 해석하는 것이 적절하다.
온도 제어 측면에서도 TGG는 고온기에 유입 공기 자체의 온도가 이미 높아져 외기 대비 더 높은 수준의 온도 처리를 안정적으로 확대하기 어렵다. Okada et al.(2000)가 제시한 사전 공조형 설계는 유입부 온도를 외기 대비 최대 10°C 낮춤으로써 이를 부분적으로 해소하였으나, 이러한 설계가 TGG에 일반적으로 적용되고 있는 것은 아니다. 운영 측면에서도 환기 제어, 보조 가열, 환경 계측 장치 등의 설비가 필요하며, 반복 확보를 위해서는 다수의 시설 설치가 요구되므로 비용과 설계 측면의 제약이 따른다(Rawson et al. 1995; Sinclair et al. 1995). 이러한 TGG의 구조적·운영적 특성은 SPAR, FACE, OTC 등 주요 기후변화 실험시설과의 비교를 통해 보다 분명하게 드러난다(Table 3).
Table 3
Comparison of major features among sunlit climate-change research facilities. Summarized from Horie et al.(1995), Rawson et al.(1995), Sinclair et al.(1995), Reddy et al.(2001), Morales et al.(2014), Allen et al.(2020), and Seo et al.(2021)
SPAR은 온도와 CO2 농도를 정밀하게 제어할 수 있고, 군락 수준의 광합성과 증산을 연속적으로 계측할 수 있어 기초 생리 반응의 정량화에 강점을 가진다(Reddy et al. 2001; Allen et al. 2020). 그러나 소형 밀폐 구조에 따른 인공 배지 사용과 높은 냉수 설비 비용은 군락 규모의 포장 조건 재현과 대규모 비교 실험의 적용을 제한한다.
FACE는 완전 개방 조건에서 자연광, 자연 기류, 자연토양을 유지할 수 있어 포장 재현성과 생태계 규모 연구에 유리하다. 그러나 바람 의존적 CO2 분배에 따른 농도 변동, 온도의 독립적 정밀 제어의 어려움, 그리고 높은 운영비는 주요 제약으로 작용한다(Allen et al. 2020).
OTC는 구조가 단순하고 구축 비용이 상대적으로 낮으며, 자연광과 자연토양을 유지할 수 있다는 장점을 지닌다. 그러나 온도 상승은 시설 내 공기 체류에 따른 부수적 효과로 발생하므로 처리 수준이 단일하고, 전기 저항열 가열 방식으로는 일변화에 따른 자연스러운 온도 변화를 재현하기 어렵다. 또한 처리구 규모가 중소형에 한정되어 군락 수준의 기체교환 산정이나 다수의 연속 온도 수준을 동시에 비교하는 데에는 불리하다(Allen et al. 2020).
종합하면, TGG는 FACE의 높은 포장 재현성과 SPAR의 높은 제어 정밀성 사이에 위치하는 중간형 연구 시설으로 해석할 수 있다. TGG는 자연광 기반의 포장 유사성을 유지하면서 단일 시설 내에서 다수의 연속적 온도 수준을 동시에 구현할 수 있다. 이러한 특성은 온도 반응 곡선 도출, 생육 단계별 민감 시기 비교, 그리고 작물 모형의 반응 함수 매개변수화에 고유한 강점을 지닌다(Morales et al. 2014; Allen et al. 2020). 반면 습도와 CO2를 온도와 독립적으로 제어하기 어렵고, 미기상 불균일성과 반복성 확보의 제약이 남아 있으므로, 복합 환경 요인의 기작 규명에는 SPAR이나 FACE와의 상호보완적 활용이 요구된다.
온도구배하우스 이용 기후변화 대응 작물 연구 전망
TGG는 열환경 제어의 불균일성과 포장 재현성의 차이, 운영적 제약이 잔존하지만, 자연광 조건에서 연속적인 온도 구배를 형성하여 작물의 단계-특이적 온도 민감성과 비선형적 임계 반응을 규명할 수 있는 실험적 강점을 지닌다. 향후 연구는 이러한 현상 규명을 토대로, 한계를 보완하면서 연구 활용 범위를 확장하는 방향으로 전개될 필요가 있다.
TGG에서 축적된 연속적 온도 반응 데이터는 DSSAT, APSIM 등 작물 모델의 온도 반응 함수와 관련 매개변수를 보정하는 데 활용될 수 있다. 이들 모델에서는 발육 속도, 엽면적 확장, 광합성 및 등숙 과정을 기본 온도, 최적 온도, 상한 온도에 기반한 온도 함수와 적산 온도를 이용하여 모의하며(Jones et al. 2003; Keating et al. 2003), 이러한 온도 반응 계수의 불확실성은 모델 간 예측 편차의 주요 원인 중 하나로 보고되고 있다(Rosenzweig et al. 2014). 본 논문에서 검토한 바와 같이, 벼의 개화기 고온에 의한 화분 임성 저하(Oh et al. 2023), 콩의 임계 온도 초과 시 종자수 급감과 수확 지수 저하(Tacarindua et al. 2013) 등은 이러한 모델의 생식 생장 반응 함수를 실측 기반으로 보정하는 데 핵심적인 기초 자료가 될 수 있다.
미래 기후 조건에서 온도 상승은 수분 스트레스, 대기 CO2 농도 변화와 복합적으로 작용할 가능성이 높으므로, TGG의 온도 구배에 관수 수준이나 CO2 농도 처리를 조합하는 복합 환경 실험의 확대가 요구된다. Liu et al.(2025)은 TGG 내에서 4단계 온도 구배와 2수준 관수 처리를 조합하여 콩의 복합 스트레스 반응을 분석하였으며, 복합 처리구에서 지상부 건물중이 대조구 대비 60% 이상 감소하는 비가산적 상호작용을 보고하였다. 이러한 복합 환경 실험의 확대는 앞 절에서 언급한 수증기압포차 동반 변동의 영향을 실험적으로 분리하는 데에도 기여할 수 있다.
다만 Table 3에서 확인된 바와 같이 TGG는 습도와 CO2의 독립 제어에 한계가 있다. 이를 보완하기 위해 TGG에서 도출된 온도 반응 곡선을 SPAR의 정밀 제어 실험으로 검증하거나, FACE의 포장 규모 관측 자료와 교차 비교하는 시설 간 연계가 필요하다. 또한 TGG 내부는 피복재에 의해 자연광이 감소하고, 환기 방식에 따라 내부 기류가 포장과 다르므로(Horie et al. 1995; Morales et al. 2014), 야외 관측 자료와의 비교를 통해 이러한 시설 내 환경 차이가 작물 반응에 미치는 영향을 평가하는 체계를 갖추는 것이 필요하다.
이러한 연구의 축적은 TGG가 기후변화 조건에서의 작물 생산 취약성 평가와 적응 전략 수립에 실증적 근거를 제공하는 연구 시설로 발전하는 데 기여할 것으로 판단된다.
