©The Korean Society for Bio-Environment Control, All rights reserved.
ABSTRACT
서 론
재료 및 방법
1. 노지재배 품종별 동해 조사
2. 수령별 저온 수준별 피해 발생
결과 및 고찰
1. 노지재배 품종별 동해
2. 수령 및 저온 수준별 동해
서 론
올리브(Olea europaea L.)는 물푸레나무(목서)과에 속하는 상록교목으로 기원전 2,000-3,000년에 시리아, 이스라엘을 중심으로 중동 일대에서 재배 및 채유가 시작되었다(Zohary와 Spiegel-Roy, 1975). 올리브는 더 나은 양과 품질의 오일에 대한 수요 증가로 중동 지역에서 고위도로 재배면적이 점점 확대되어, 현재는 북위 30°에서 45° 사이 지중해 지역에서 가장 중요한 상록과수 중 하나가 되었다(Candev 등, 2009; Mancuso, 2000). 다른 아열대 과수에 비해 올리브는 비교적 저온에 잘 견디지만, 고위도 재배지에서는 겨울과 이른 봄의 최저온도가 생산성과 품질의 중요한 제한 요인이 되어 왔다(Aybar 등, 2015; Sanzani 등, 2012; Yang 등, 2005). 내한성이 약한 올리브 품종은 겨울 기온이 -7°C 이하로 낮아지면 나무의 윗부분부터 손상되어(Vitagliano와 Sebastiani, 2002), 수확량 감소로 이어지거나 심하면 고사에 이를 수도 있다(Palliotti과 Bongi, 1996). -12°C 이하의 온도에서는 대부분의 품종들이 심각한 동해를 받지만(Gomez Del Campo와 Barranco, 2005; Rahemi 등, 2016), 일부 품종은 -12°C에서 -18°C 사이의 온도에서도 저온에 대한 내성을 나타내기도 한다(Fiorino와 Mancuso, 2000). 따라서 고위도의 재배지 확장에서 내한성이 강한 품종을 선택하여 심는 것이 재배의 기본이라 할 수 있다.
올리브의 재배지역 확대에 따라 내한성에 관한 연구가 다양하게 이루어지고 있는데 저온에 따른 수체 내 전기전도도, 호르몬, methane dicarboxylic aldehyde(MDA), superoxide dismutase(SOD) 함량 변화 등이 보고되었다(Cansev 등, 2011; Guo 등, 2004; Hashempour 등, 2014; Qin 등, 2011). 기공 밀도(Roselli 등, 1989), 이온 누출(Barranco 등, 2005), 탄수화물 및 전분 농도(Lavee, 1986)와 같은 식물 생리학적 특성을 지표로 활용하여 올리브의 저온 내성 유전자형을 선발하는 데 사용되고 있다. 동해는 온도 수준과 지속시간, 작물의 생육단계, 재배품종, 재배지의 지형과 토양특성에 따라 다르게 나타날 수 있다(Choi와 Lee, 1976; Gomez Del Campo와 Barranco, 2005; Kim 등, 2007).
주요 올리브 재배 국가들보다 기온이 낮은 우리나라의 경우도 내한성이 재배의 주요 제한요인이 되지만 제주지역에서는 올리브의 정상적인 영양생장 및 착과가 가능한 것으로 보고되었다(Lim 등, 2018). 경남의 남부지역의 경우 겨울 최저기온이 -10°C 이하로 떨어지는 경우가 빈번하여 경제적 재배가 어려운 것으로 여겨져 왔다. 그러나 지구온난화가 지속되고 있고 농업기술센터나 농업인은 새로운 소득작물에 대한 관심이 높아 수년 전부터 시범적으로 재배를 시도하고 있으나 정확한 품종 정보 없이 재식하여 동해를 심하게 받거나 착과가 제대로 되지 않은 경우가 많다.
본 연구는 경남 해안지역에서 내한성이 강한 올리브 품종을 선발하여 노지재배 가능성을 검토하고, 저온 수준별 피해 정도를 파악하여 재배관리의 기초자료를 도출하고자 수행하였다.
재료 및 방법
1. 노지재배 품종별 동해 조사
경남 해안지역에서 내한성이 강한 품종을 선발하여 노지재배 가능성을 평가하기 위하여 거제시 거제면에 위치한 농업기술센터 시험포(34°51'19"N 128°34'50"E)와 남해군 이동면에 위치한 농업기술센터 시험포(34°48'58"N 127°55'19"E)에서 시험을 수행하였다. 2021년 4월에 국립원예특작과학원에서 유망품종으로 선발했던 올리브 ‘Frantoio’, ‘Verdale’, ‘Maurino’, ‘Leccino’, ‘Koroneiki’, ‘Arbequina’ 품종을 열간거리 2.5-3m, 주간거리 1-1.5m로 5주씩 재식하였다. 거제 포장의 토양은 사질의 마사토, 남해 포장은 양토였으며, 두 포장 모두 물빠짐이 양호하였다. 2022년 12월부터 2023년 1월 동안에 남부지방에 한파가 심하여 올리브 나무에 동해가 다수 발생하였다. 각 농업기술센터의 농업기상관측장비에서 측정한 기온 자료를 조사한 결과 2022년 12월 26일 거제는 -6.6℃, 남해는 -8℃로 내려갔으며, 2023년 1월 25일에는 거제 -10.9℃, 남해 -12.3℃까지 하강하였다. 2023년 3월 20일에 품종별로 잎이 고사한 가지를 피해로 간주하고 그 비율을 평균값과 표준오차로 표시하였다.
2. 수령별 저온 수준별 피해 발생
2024년 5월에는 2022-2023년에 발생한 동해 결과 내한성이 강하다고 판단되었던 Arbequina 품종의 2, 3, 4년생 묘목을 사용하여 수령별 내동성 차이를 조사하였다. 모든 식물체는 일정한 크기의 묘목을 선정하여 시판 원예용 상토(Baroker, Seoul Bio, Eumseng)를 담은 플라스틱 포트(W 20cm × L 15cm × H 21cm)에 옮겨 심은 후, 2월 10일부터 4월 15일까지 비가림 온실에 3개월간 순화시켰다.
저온처리는 수령별 생육이 비슷한 나무를 5주씩을 골라 처리 전 24시간 동안 상온에서 순화 처리하여 4℃에서 2시간 동안 예비 저온처리 후 실험에 사용하였다. 냉각 속도가 시간당 8-15℃를 넘으면 내한성이 과대 평가될 수 있기 때문에(Probsting과 Sakai, 1979), 저온처리는 저온챔버(SH-75BS, Seyoung Scientific Co., Korea)에서 20분당 1℃씩 낮추었다. 상온(25℃)을 대조구로 하였으며, 최저온도 -5℃~-14 ℃까지 7수준으로 나누어 목표 온도 도달 10분 후 꺼내어 3시간 후 잎의 변색 및 말림 개수를 세어 육안에 의한 피해율을 산출하였다. 실험 묘목의 가지는 전해질 누출률과 TTC 검정에 사용하였다.
전기전도도 측정법은 저온으로 세포막이 손상되어 누출되는 전해질 비율을 측정하는 방법(Maier 등, 1994)으로 본연구에서는 Kim 등(2007)이 사용한 방법으로 조사하였다. 각 처리의 5주 묘목으로부터 직경 1.5-2.2mm 1년생 가지를 채취하여 눈이 포함되지 않은 마디 중간부분을 5g으로 정량하여 40mL 증류수에 담근 후 20℃에서 15시간 배양하고 배양한 용액을 전기전도계(pH/LF 12, Schott, Germany)로 전해질 누출량(C1)을 측정하였다. 각 시료액은 95℃에서 30분간 중탕하여 전해질을 파괴시킨 후 20℃에서 15시간 배양하여 용액의 전해질 누출량(C2)을 측정하고 전해질 누출률을 (C1/C2) × 100 공식으로 계산하였다.
TTC(2.3,5-triphenyl tetrazolium chloride) 검정은 용액이 붉은색의 triphenylformazan으로 환원되는 정도로서 조직의 활력을 판정하는 방법으로 Kim 등(2007)의 방법을 따랐다. 저온 처리한 묘목에서 직경이 1.3-2.1mm인 가지의 수피를 취해 0.5g으로 정량한 후 25℃에서 0.1% TTC 용액에 15시간 동안 침지 처리를 하였다. 처리 후 수피를 증류수로 2회 수세한 후 70℃의 10mL 무수에탄올에 30분간 담근 후 붉은색의 triphenylformazan을 추출한 용액을 UV spetrophotometer (JP/UV-16501, Shimadzu Co., Kyoto, Japan)로 530nm에서 흡광도를 측정하여 무처리 대비구의 흡광도에 대한 백분율을 구하였다. 전해질 누출률과 TTC 검정 결과는 평균값과 표준오차로 표시하였다.
시험성적의 통계분석은 육안에 의한 피해율은 t-test로 비교하였고, 전해질 누출률과 TTC검정, 수령별 동해피해는 Duncan’s multiple range test 5% 수준에서 검정하였다.
결과 및 고찰
1. 노지재배 품종별 동해
2022년 12월 26일의 거제 -6.6℃, 남해 -8℃ 및 2023년 1월 25일의 거제 -10.9℃, 남해 -12.3℃의 최저기온을 경과한 4년생 올리브 6품종의 육안 판별에 의한 피해 정도는 Fig. 1과 같다. 대부분의 품종들이 12월의 저온에서는 일부 잎의 갈변 및 말림으로 가벼운 피해를 받았다가 이듬해 1월의 저온에서 주로 피해를 받은 것으로 관찰되었다. 품종별 피해율을 비교해 보면, 거제에는 ‘Arbequina’, ‘Verdale’ > ‘Maurino’ > ‘Frantoio’ > ‘Leccino’ > ‘Koroneiki’ 순으로, 남해에서는 ‘Arbequina’ > ‘Verdale’ > ‘Leccino’ > ‘Frantoio’ > ‘Maurino’ > ‘Koroneiki’ 순으로 동해에 강한 것으로 나타났다. ‘Arbequina’ 품종은 거제에서는 피해가 없었고, 남해에서는 피해율이 17%로 다른 품종보다 낮아 내한성이 강한 반면, Koroneiki는 두 시험포 모두 피해율이 90% 이상으로 내한성이 약한 것으로 판단되었다. 이러한 결과는 Arbequina 품종이 내한성이 비교적 강하여 -10.6℃에서도 견디지만(Gomez Del Campo와 Barranco, 2005; Ruiz 등 2006), ‘Koroneki’ 품종은 -4.9 ~-6.2℃에서 줄기 피해를 받고, -4.2~-4.7℃에서도 잎이 피해를 받아 저온에 약하다는 선행 연구결과(Wang 등, 2018)와 유사하였다. ‘Verdale’, ‘Maurino’, ‘Leccino’는 두 시험포 간에 피해율 차이가 컸는데, 이는 동해 당시의 재배지 기온이 달랐고, 지형과 토양특성 등에 따라 내한성이 달라지기 때문(Choi와 Lee, 1976; Gomez Del Campo와 Barranco, 2005; Kim등, 2007)으로 여겨진다. 종합하면 ‘Arbequina’ 품종이 -12.3℃의 기온에서도 1년생 가지의 피해율이 20% 이하로 나무의 생존에 큰 영향이 없었기 때문에 최저기온 -10℃ 이상인 지역에서는 안정적인 재배 가능성이 높은 것으로 판단되었다.
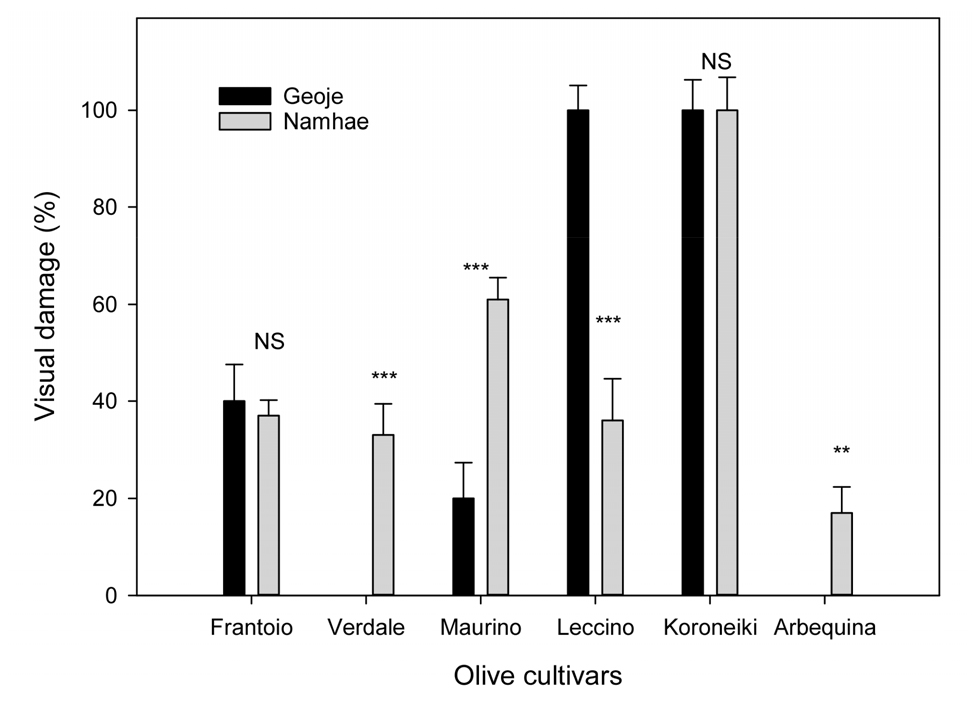
Fig. 1.
The visual cold damage on shoots in different olive cultivars after passing the lowest temperature, -10.9℃ in Geoje and -12.3℃ in Namhae in January, 2023. Vertical bars are SE (n = 5). Values sharing the same letter at each temperature are not significantly different from each other by least significant difference at p ≤ 0.05.
2. 수령 및 저온 수준별 동해
저온챔버에서 수령이 다른 ‘Arbequina’ 품종을 저온 수준을 달리하여 처리한 결과 육안으로 판별한 피해율은 Fig. 2와 같다. 2년생 나무의 동해 발생률은 -5℃에서는 5.7%로 생육에 큰 지장이 없는 수준이었으며 -10℃에서 28%, -14℃에서는 78%까지 온도가 하강함에 따라 점진적으로 증가하였다. 3년생은 -10℃까지 하강하더라도 피해율이 8.6%로 2년생의 절반 이하였으나 -11℃에서는 23%으로 급격히 증가하였고, -14℃에서는 55%였다. 반면 4년생의 피해율은 -10℃까지는 3년생과 비슷하였고, -13℃까지도 12.3%로 높지 않았으나 -14℃에서 45.2%로 증가하였다. 종합하면 20% 이상 피해를 받는 온도는 2년생은 -10℃, 3년생은 -11℃, 4년생은 -13~-14℃로 수령별 피해를 받는 온도가 다름을 알 수 있었다. 앞의 시험 1 결과에서 거제지역 4년생 ‘Arbequina’가 -10.9℃에서 동해피해를 받지 않았고 남해지역에서는 -12.3℃에서 17% 동해피해를 받은 것을 감안하면, 저온챔버에는 4년생 피해율과 차이가 있는 것은 저온처리 전후의 온도, 습도, 토양수분 등의 외부 환경조건(Choi와 Lee, 1976) 및 식물체의 휴면심도(Song 등, 2000), 저장양분(Choi 등, 2005) 때문으로 판단된다.
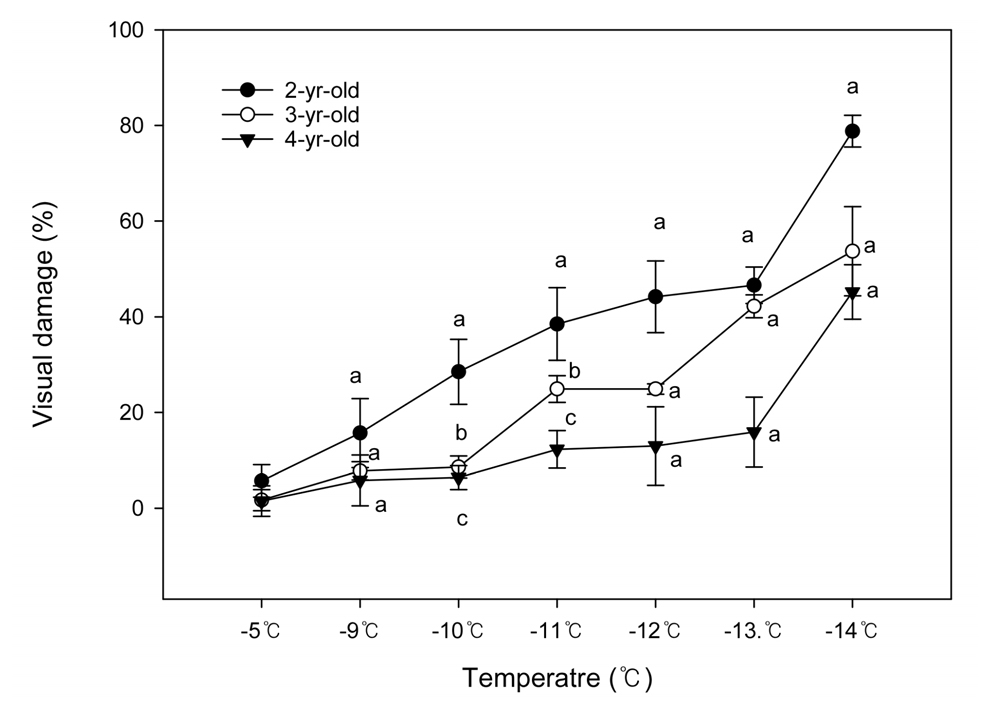
Fig. 2.
The visual cold damage on shoots of differently aged ‘Arbequina’ olives under low-temperature treatment. Vertical bars are SE (n = 5). Values sharing the same letter at each temperature are not significantly different from each other by Duncan’s multiple range test at p ≤ 0.05.
육안에 의한 동해 판별은 관찰자의 경험적인 판단을 하기 때문에 객관적 수치화에 어려움이 있으므로 전해질 누출률과 TTC 용액 처리 후 흡광도를 측정하여 검정한 결과는 Fig. 3과 Fig. 4와 같다. 전해질 누출률은 온도가 낮고 수령이 어릴수록 높은 경향이었다(Fig. 3). 2년생은 -5℃에서 77%, -11℃에서 83%, -14℃에서 88%를 보인 반면, 4년생은 -5℃에서 58.6%, -11℃에서는 71.3%, -14℃에서는 78%로 그 비율이 낮아졌다. 이러한 결과는 최저온도가 낮을수록, 유목의 수령이 어릴수록 동해가 증가한 시험 1의 결과와 유사하였다. 한편, ‘Arbequina’ 품종의 전해질 누출률은 -11.8℃에서 증가한다는 선행 연구결과(Barranco 등, 2005)와는 차이가 있었다.
조직활력 정도를 나타내는 TTC 검정에 의한 흡광률은 그 값이 낮을수록 동해가 심하다고 판단할 수 있는데(Kim 등, 2007), 본 시험에서 처리온도가 낮아질수록 흡광률이 감소하여 동해가 증가하였음을 확인하였다(Fig. 4). 흡광률은 -5℃에서 84.5-94.2%를 보였고, -11℃까지 감소하는 경향이었으나 수령별 차이는 보이지 않았다. 그러나 처리온도가 -14℃로 낮아졌을 때 2년생 52%, 3년생 66.4%, 4년생 69.5%로 2년생 가지에서 크게 감소하고, 3년생과 4년생 간에는 뚜렷한 차이가 없었다.
연구결과를 종합하면, 거제와 남해에서 각각 1월의 최저기온 -10.9℃와 -12.3℃ 에 각각 노출된 후의 내한성은 ‘Arbequina’ 품종이 가장 강하고, ‘Koroneiki’가 가장 약한 것으로 나타나 겨울 최저기온이 -10℃ 이상인 지역에서 ‘Arbequina’ 재배 가능성이 높은 것으로 판단되었다. 2-4년생의 수령이 다른 ‘Arbequina’ 묘목의 저온 수준별 피해율, 전해질 누출률, TTC 검정을 통한 흡광률을 조사한 결과, 4년생이 2년생보다 높은 내한성을 가진 것으로 확인되었다. 따라서 ‘Arbequina’ 묘목을 재식하더라도 나무가 어릴수록 동해를 받지 않도록 겨울철 보온관리가 중요함을 알 수 있었다.
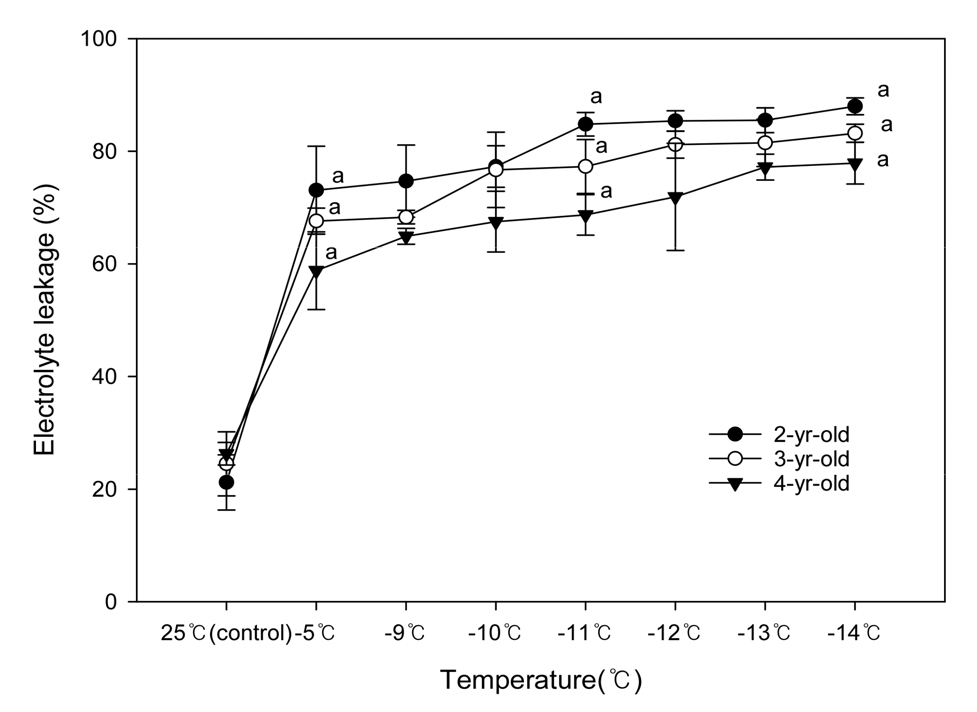
Fig. 3.
Electrolyte leakage from shoots of differently aged ‘Arbequina’ olives after low-temperature treatments. Vertical bars are SE (n = 5). Values sharing the same letter at each temperature are not significantly different from each other by Duncan’s multiple range test at p ≤ 0.05.
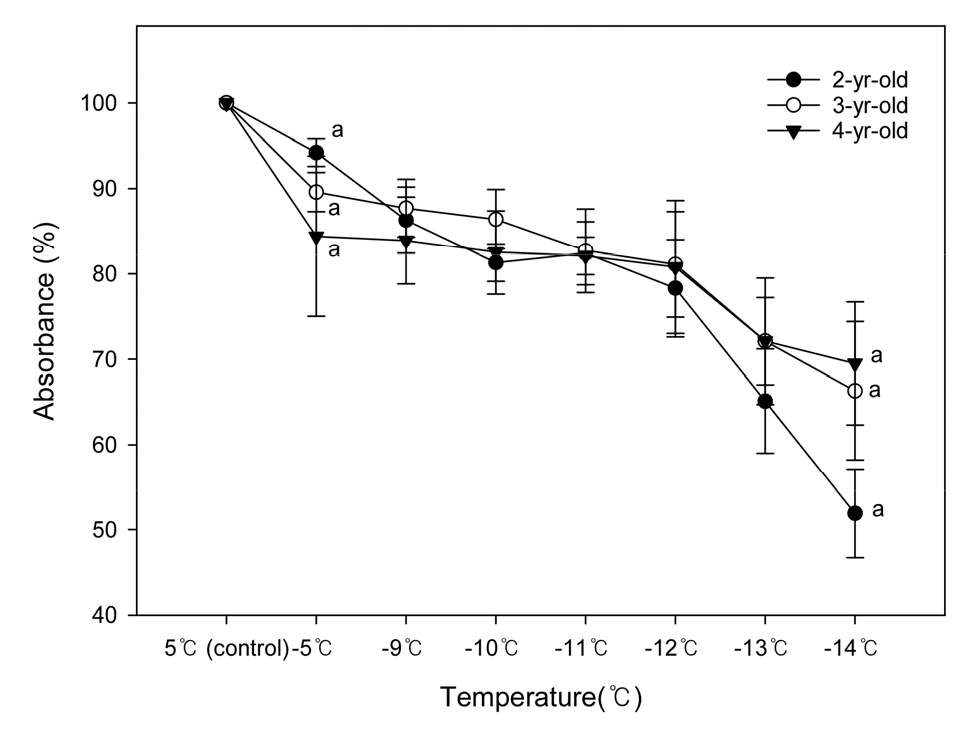
Fig. 4.
Absorbance by the TTC test from shoots of differently aged ‘Arbequina’ olives after low-temperature treatments. Vertical bars are SE (n = 5). Values sharing the same letter at each temperature are not significantly different from each other by Duncan’s multiple range test at p ≤ 0.05.
Acknowledgements
본 연구는 농촌진흥청 공동연구과제(과제번호: PJ016091032021)의 지원에 의해 수행되었음.
References
Aybar V.E., J.P.D. Melo-Abreu, P.S. Searles, A.C. Matias, C. Del Río, J.M. Caballero, and M.C. Rousseaux 2015, Evaluation of olive flowering at low latitude sites in Argentina using a chilling requirement model. Spanish J Agric Research 13:10. doi:10.5424/sjar/2015131-6375
10.5424/sjar/2015131-6375Barranco D., N. Ruiz, and M. Gomez-del Campo 2005, Frost tolerance of eight olive cultivars. Hort Sci 40:555-560. doi:10.21273/HORTSCI.40.3.558
10.21273/HORTSCI.40.3.558Candev A., H. Gulen, and A. Eris 2009, Cold-hardiness of Olive (Olea europaea L.) Cultivars in Cold Acclimated and Non-acclimated Stages: Seasonal Alternation of Antioxidative Enzymes and Dehydrin-Like Proteins. J Agric Sci 147:51-61. doi:10.1017/S0021859608008058
10.1017/S0021859608008058Cansev A., H. Gulen, and A. Eris 2011, The activities of catalase andascorbate peroxidase in olive (Olea europaea L. cv. Gemlik) under low temperature stress. Hortic Environ Biotechnol 52:113-120. doi:10.1007/s13580-011-0126-4
10.1007/s13580-011-0126-4Choi H.O., and J.H. Lee 1976, Studies on low temperature injury at each growth stage in rice plant. Korean J Crop Sci 21:203-210.
Choi S.T., S.M. Kang, D.S. Park, Y.W. Yoon, and G.H. Ahn 2005, Tree responses of 'Fuyu' persimmon to different degrees of early defoliation on fruit characteristics at harvest and tree development the next season. Hortic Environ Biotech 46:136-139.
Fiorino P., and S. Mancuso 2000, Differential thermal analysis, deep supercooling and cell viability in organs of Olea europaea at subzero temperatures. Advances in Horticultural Sci 14:23-27.
Gomez Del Campo M., and D. Barranco 2005, Field evaluation of frost tolerance in 10 olive cultivars. Plant Gen Res 3:385-390. doi:10.1079/PGR200592
10.1079/PGR200592Guo H., S. Gao, F. Zhao, and F. Li 2004. Effect of cold acclimation on several enzyme activities in Euonymus radicans 'emorald & gold' and its relation to semi-lethal temperature. Forest Stud China 6:10-17.
10.1007/s11632-004-0002-8Hashempour A., M. Ghasemnezhad, R. Fotouhi, and M.M. Ghazvini Sohani 2014, Olive (Olea europaea L.) freezing tolerance related to antioxidant enzymes activity during cold acclimation and acclimation. Acta Physiol Plant 36:3231-3241. doi:10.1007/s11738-014-1689-3
10.1007/s11738-014-1689-3Kim H.C., K.S. Bae, J.H. Bae, and T.C. Kim 2007, Freezing hardiness according to dormancy level and low temperature in persimmon (Diospyros kaki). J Bio-Environ Cont 16:269-273.
Lavee S. 1986, Involvement of plant growth regulators and endogenous growth substances in the control of alternate bearing. Acta Hort 239:311-322. doi:10.17660/actahortic.1989.239.50
10.17660/ActaHortic.1989.239.50Lim C.K., H.J. An, and J.H. Noh 2018, Vegetative Growth and Fruit Characteristics of Introduced Olive (Olea europaea L.) Varieties Cultivated in Jeju Island, Korea. J Korean Soc International Agric 30:113-119. doi:10.12719/KSIA.2018.30.2.113
10.12719/KSIA.2018.30.2.113Maier F.P., N.S. Lang, and J.D. Fry 1994, Evaluation of an electrolyte leakage technique to predict St. Augustinegrass freezing tolerance. J Am Soc Hortic Sci 29:316-318. doi:10.21273/HORTSCI.29.4.316
10.21273/HORTSCI.29.4.316Mancuso 2000, Electrical resistance changes during exposure to low temperature measure chilling and freezing tolerance in olive tree (Olea europea L.) plants. Plant Cell and Environ 23:291-299. doi:10.1046/j.1365-3040.2000.00540.x
10.1046/j.1365-3040.2000.00540.xPalliotti A., and G. Bongi 1996, Freezing Injury in the Olive Leaf and Effects Mefluidide Treatments. Hort Sci 71:57-63. doi:10.1080/14620316.1996.11515382
10.1080/14620316.1996.11515382Probsting E., and A. Sakai 1979, Determining T50 of peach flower buds with exotherm analysis. J Am Soc Hortic Sci 14:597-598. doi:10.21273/HORTSCI.14.5.597
10.21273/HORTSCI.14.5.597Qin X., Y. Liu, S. Mao, T. Li, H. Wu, C. Chu, and Y. Wang 2011, Genetic transformation of lipid transfer protein encoding gene in Phalaenopsis amabilis to enhance cold resistance. Euphytica 177:33-43. doi:10.1007/s10681-010-0246-4
10.1007/s10681-010-0246-4Rahemi M., F. Yazdani, S. Sedaghat 2016, Evaluation of freezing tolerance in olive cultivars by stomatal density and freezing stress. Int J Hortic Sci Technol 3:145-153. doi:10.22059/ijhst.2016.62914
Roselli G., G. Benelli, and D. Morelli 1989, Relationship Between Stomatal Density and Winter Hardiness in Olive (Olea europaes L). Hort Sci 64:199-203. doi:10.1080/14620316.1989.11515945
10.1080/14620316.1989.11515945Ruiz N., D. Barranco, and H.F. Rapoport 2006, Anatomical response of olive (Oleaeuropaea L.) to freezing temperatures. J Hortic Sci Biotech 81:783-790. doi:10.1080/14620316.2006.11512138
10.1080/14620316.2006.11512138Sanzani S.M., L. Schena, F. Nigro, V. Sergeeva, A. Ippolito, and M.G. Salerno 2012, Abiotic diseases of olive. J Plant Pathology 94:469-491. doi:10.4454/JPP.FA.2012.069
Song G.C., I.M. Choi, and M.D. Cho 2000, Cold hardiness in relation to vine management in 'Cambell Early' grapevines. Kor J Hort Sci Technol 18:387-390.
Vitagliano C., and L. Sebastiani 2002, Physiological and biochemical remarks on environmental stress in olive (Olea europaea L.). Acta Horticulturae 586:435-440. doi:10.17660/ActaHortic.2002.586.89
10.17660/ActaHortic.2002.586.89Wang J., L. Ma, M. Gómez-del-Campo, D. Zhang, Y. Deng, and Z. Jia 2018, Youth tree behavior of olive (Olea europaea L.) cultivars in Wudu, China: cold and drought resistance, growth, fruit production, and oil quality. Sci Hort 236: 106-122. doi:10.1016/j.scienta.2018.03.033
10.1016/j.scienta.2018.03.033Yang M.T., S.L. Chen, C.Y. Lin, and Y.M. Chen 2005, Chilling stress suppresses chloroplast development and nuclear gene expression in leaves of mung bean seedlings. Planta 221:374-385. doi:10.1007/s00425-004-1451-y
10.1007/s00425-004-1451-y15599759Zohary M., P. Spiegel-Roy 1975, Beginnings of fruit growing in the old world. Science 187:319-327. doi:10.1126/science.187.4174.319
10.1126/science.187.4174.31917814259

