

서 론
최근 한반도의 기상조건은 장마철 강우, 일조 부족 및 변색기(7~8월) 고온으로 인해 포도의 착색이 불량함과 동 시에 포도를 비롯한 낙엽과수의 동해 발생 또한 증가하고 있어 착색과 내한성이 우수한 포도 품종에 대한 요구가 증가하고 있다(Kwon 등, 2011; Yamane 와 Shibayama, 2006; Shim 등, 2004).
개량머루는 착색이 우수하고, 내한성과 내병성이 우수 한 것으로 알려져 우수한 포도 품종으로 주목을 받고 있으며, 재배면적은 250~300ha로 추정되고 있고 가공량 은 2009년 1,118톤으로 파주에서만 연간 300톤이 생산 되고 있다(MFAFF, 2009). 또한 항산화 능력이 우수한 anthocyanin과 t-resveratrol와 같은 phenolic compound의 함량이 높아 가공용 품종으로 주목 받고 있다(Lee 등, 2008). 그러나 개량머루에 관한 연구는 미비한 실정이며 육성과정과 양친이 불분명하고 기존 재배 품종과의 유연 관계가 분명하게 구명되지 못하고 있다(Park 등, 2003; Park과 Lee, 2007). 특히 생육 특성과 재배 기술 등에 관한 연구가 거의 없어 품종 특성에 맞는 재배 기술 개 발을 위해 적정 수확시기, 성숙기 과실 특성과 같은 기 초 자료가 지속적으로 요구되고 있다.
식물체의 정확한 구조와 각 기관의 조직 발생 및 발달 과정을 파악하는 것은 식물체를 이용하는데 있어 매우 중요한 과정이다(Cho 등, 2000). 과실의 해부학적 관찰을 통해 세포분열 정지기는 후지 사과가 만개 후 35일 전후 이고 유명 복숭아는 만개 후 40~47일이며 Sauvignon Blanc 포도는 28일 이전인 것이 구명되었다(Cho 등, 2000; Hardie 등, 1996; Park과 Park, 2000) 또한 생리장 해의 발생 원인 구명 및 재배 기술 개발에 이용하여 배 의 석세포와 동녹의 발생시기와 원인 및 복숭아의 바람 들이 발생 원인을 구명하였다(Cho 등, 2000; Choi 등, 2003; Kim 등 2003; Seo와 Park, 2003). 또한 기온에 따른 사과의 과피층 형성과 포도에서 탄닌 세포의 기능, 과립의 외부 유관속의 발달 과정, 소립과 발생 원인 및 화진현상의 형태적 차이 등이 해부학적 연구를 통하여 구명하였다(Chatelet 등, 2008; Fernandez 등, 2006; Fougére-Rifot 등, 1993; Fougére-Rifot 등, 1995). 이를 위해서는 만개기 이후 과실의 해부학적 특성이 구명되어 야 하지만 국내에서의 연구는 매우 미비한 실정이다.
따라서 본 연구는 개량머루의 생육기별 액아의 발달 과정, 과실 품질의 변화를 관찰하고 과실 조직을 해부학 적 관찰하여 개량머루의 특성과 생리를 이해하기 위한 기초 자료로 이용하고자 하였다.
재료 및 방법
경기도 안성에 위치한 농가에서 2006년에 재식거리 3.6m × 1.8m의 덕식으로 재배중인 개량머루를 이용하였다. 개량머루 액아는 6월부터 10월까지 1개월 간격으로, 액아 를 종축으로 절단하여 실체현미경(BX-51, Japan)을 이용 해 관찰한 후, 괴사 형태를 Kim 등(2011)의 방법으로 주 아괴사, 부아괴사, 눈괴사 및 정상눈으로 구분하였다.
시기별 과실 품질은 개화기부터 수확기까지 1주 간격 으로 과실을 수확하여 과방중, 과립중, 종경 및 횡경을 측정하였으며, 착즙한 과즙을 이용해 당도를 측정하고 적정산도는 과즙 10mL과 증류수 40mL을 혼합한 후 0.1N NaOH를 첨가하여 pH 8.1에 도달되었을 때의 첨 가량을 주석산 상당량으로 환산하였다.
유리당과 유기산 분석을 위해 과즙을 12,000rpm에서 5 분간 원심분리하고 상등액 1mL를 증류수로 10배 희석하 여 0.45μm syringe filter(Agilent, US)로 여과하였다. HPLC(Agilent, 1200 series, USA)는 ZORBAX carbohydrate 컬럼(150 × 4.6mm I.D; particle size 5.0μm)과 refractive index detector(RID)를 이용하여 검출하였다. 유기 산은 Allure organic acids 컬럼(150 × 4.6mm I.D; particle size 5.0μm)과 diode array detector(DAD)를 이용하여 210nm에서 검출하였다. 시료의 개별 유리당과 유기산은 표준물질(Chromadex, USA)과의 retention time, spectrum을 비교하고 spike test를 실시하여 동정하였다.
개량머루 과피 조직의 형태적 특성을 관찰하기 위해 과립의 적도부위에서 과피절편을 채취하였다. 세포 고정 의 모든 과정은 Park 등(2013)의 방법과 같이 2.5% glutaraldehyde 용액과 1% osmium tetroxide 용액에 세포 를 고정시키고 ethyl alcohol의 농도를 단계별로 높여 탈 수시킨 후 조직에 epon을 침투시키기 위하여 침지하여 캡슐에 매몰하였다. 조직의 절편을 만들기 위해 캡슐을 ultramicrotome(PT-X, RMC, USA)으로 잘라 PAS 염색하 여 광학현미경(BX51, Olympus, Japan)으로 관찰하였다.
결 과
개량머루 신초에 발생된 액아의 특성을 구명하기 위하 여 6월부터 1개월 간격으로 액아의 절단면 형태와 괴사 발생을 조사하였다. 개량머루의 액아는 종축으로 절단하 였을 때 1개의 주아를 중심으로 좌우에 부아가 1개씩 위치하는 형태였다(Fig. 1). 8월 이후에는 엽원기와 마디 의 형태가 육안으로 구분이 가능하고 화원기가 쉽게 관 찰되었다. 또한 주아와 부아를 각각 감싸고 있는 털의 구분이 가능하였다.
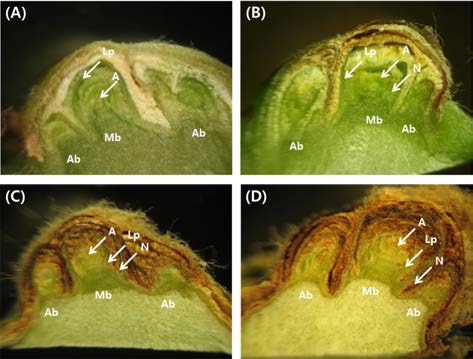
시기에 따른 눈의 크기와 괴사 발생을 조사한 결과 (Table 1), 액아의 크기는 6월(3.4mm)부터 10월(4.1mm)까 지 계속 증가했다. 6월에서 10월에는 정상눈이 98.0~100% 로 주아와 부사의 괴사가 전혀 발생하지 않았다. 마디 위 치에 따른 액아의 특성을 조사한 결과(Table 2) 크기는 3.3~5.0mm이었고 신초의 3~5번과 10번 마디에 있는 액 아가 가장 컸으며 모든 눈에서 괴사가 거의 발생하지 않았다.
Table 1.
Bud size and necrosis in Gaeryangmeoru grapes according to investigated time.
| Investigated time | Bud sizez (mm) | Normal bud (%) | Main bud necrosis (%) | Accessory bud necrosis (%) | Whole bud necrosis (%) |
|---|---|---|---|---|---|
| June | 3.4 by | 100.0 a | - | - | - |
| July | 3.2 b | 100.0a | - | - | - |
| August | 3.8 ab | 100.0a | - | - | - |
| September | 3.7 ab | 98.0a | 2.0a | - | - |
| October | 4.1 a | 100.0 a | - | - | - |
Table 2.
Bud size and necrosis in Gaeryangmeoru grapes according to bud position of the shoot.
| Bud position | Bud sizez (mm) | Normal bud (%) | Main bud necrosis (%) | Accessory bud necrosis (%) | Whole bud necrosis (%) |
|---|---|---|---|---|---|
| 1y | 4.8 abx | 100.0 a | - | - | - |
| 2 | 4.1 bcd | 96.0 a | 4.0 | - | - |
| 3 | 5.0 a | 100.0 a | - | - | - |
| 4 | 4.2 abc | 100.0 a | - | - | - |
| 5 | 4.4 abc | 100.0 a | - | - | - |
| 6 | 3.7 cd | 100.0 a | - | - | - |
| 7 | 3.7 cd | 100.0 a | - | - | - |
| 8 | 3.3 d | 100.0 a | - | - | - |
| 9 | 3.3 d | 100.0 a | - | - | - |
| 10 | 4.4 abc | 100.0 a | - | - | - |
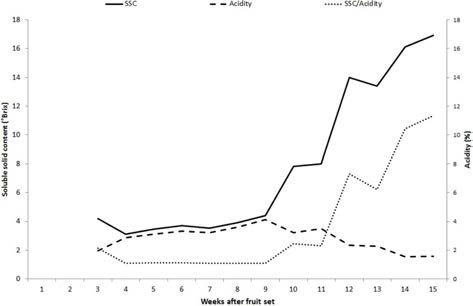
개량머루 과실의 발달 과정과 특성을 구명하기 위하여 착과 후 1주에서 수확기까지 1주일 간격으로 과실의 크 기와 품질을 조사하였다(Fig. 2). 성숙기에는 과방중 128.8g, 과립중1.2g이었으며 과립수는 98.0개 였고 과립 의 종경 및 횡경은 각각 12.2mm와 12.1mm이었으며 과 형지수는 1.01이었다. 당도와 산도는 동일한 경향의 함 량 변화를 보이다 만개 9주를 기점으로 급격하게 변화 하였으며, 성숙기에는 16.9°Brix와 1.57%로 당산비는 10.8이었다.
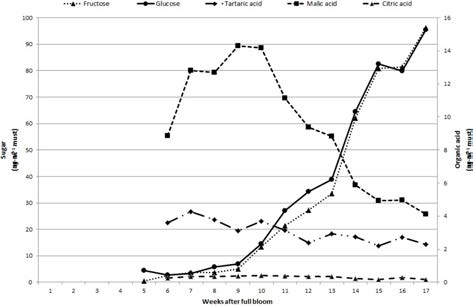
개량머루의 과즙에서는 과당과 포도당만이 검출되었 고 자당을 비롯한 다른 당은 검출되지 않았다(Fig. 3). 과당과 포도당의 비율은 만개 후 5주에는 0.09였으며 성숙기에 가까워질수록 과당의 비율이 높아져 최종 수 확기에는 1:1에 도달하였다. 과당과 포도당의 함량은 성 숙기까지 지속적으로 증가하였고 만개 9주부터 빠르게 증가하였다. 사과산과 주석산이 개량머루의 주요 유기 산이었고 그 중 사과산이 가장 많았으며 구연산은 미량 검출되었다. 사과산은 만개 후 9~10주에 가장 많고 변 색기부터 급격하게 감소하였으며 주석산은 만개 후 7주 부터 감소하였다.
주요 생육기의 과실 조직 및 세포의 형태적 특성을 광 학현미경을 이용하여 관찰하였다(Fig. 4). 개화기에는 과 실이 주변부와 중심부로 크게 나누어지며 중심부는 2개 의 심실과 각 심실내에 2개의 배주가 있어 총 4개의 배 주를 갖고 있었다. 개화기의 주변부는 유관속 조직에 의 해 비슷한 크기로 양분되고 있었으며 outer epidermis, hypodermis, inner parenchyma tissue에서 타닌이 관찰되 었으며 inner epidermis에서는 관찰되지 않았다. 배주는 외주피와 내주피의 구분이 명확하게 관찰되었다. 개화기 에서 수정까지는 약 3~4일이 소요되었으며 수정 직후에 는 유관속 조직 안쪽의 유조직에서 급격히 타닌이 사라 졌고 일부 퇴화된 배주가 수축되어 있는 것이 관찰되었 다. 개화 1주일 후에는 수정이 완료된 후에도 계속해서 퇴화되는 배주가 관찰되었다. 착과 7주 후에는 outer epidermis를 비롯한 모든 조직에서 타닌이 사라졌으며 과실내의 타닌은 변색기부터 다시 축적되는 것으로 관찰 되었다.
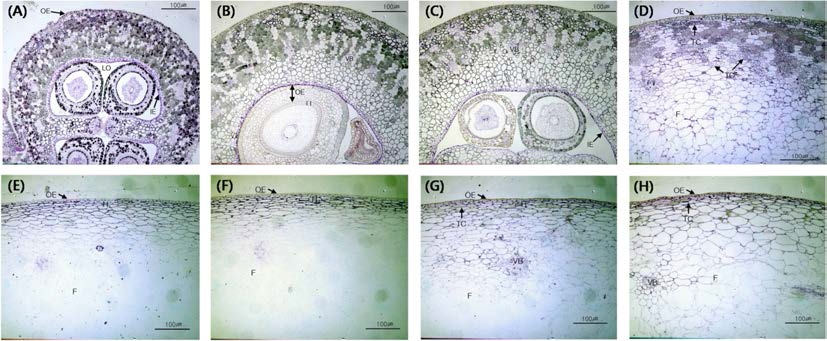
Fig. 4.
Morphological development in Gaeryangmeoru grapes. (A) Anthesis, (B) 2~3 days after anthesis, (C) 1 week after full bloom, (D) 2 weeks after full bloom. (E) 3 weeks after full bloom, (F) 6weeks after full bloom, (G) veraison, (H) maturity F, fresh; IE, inner epidermis; LO, locule; OE, outer epidermis; TC, tannin cell; VB, vascular bundle.
생육단계에 따른 과실의 비대양상을 확인하기 위하여 vascular bundle tissue에서 epidermis layer까지 세포층수와 두께의 변화를 측정하여 조사하였다(Table 3). 개화기에는 vascular bundle tissue와 epidermis 사이의 세포층은 9.3층이 었고, 만개 1주 후에는 15.3층이었으며 만개 후 2주와 성숙 기에는 19.3이었다. Vascular bundle tissue와 epidermis사이 의 두께는 60.32μm(개화기)에서 230.48 μm(성숙기)까지 증 가되었다. 성숙기 개량머루의 과피는 한 층의 epidermis 세 포와 7~10층의 hypodermis 세포로 구성되었다.
Table 3.
Cell layer numbers and thickness from epidermis layer to vascular bundle tissue at each development stage in Gaeryangmeoru berries.
| Growth Stage | Cell layer numbers between epidermis and vascular bundle | Thickness ( m) | ||
|---|---|---|---|---|
| Anthesis | 9.3 cz | 60.32 d | ||
| 1 week after full bloom | 15.3 b | 112.38 c | ||
| 2 weeks after full bloom | 18.6 a | 169.52 b | ||
| Maturity | 19.3 a | 230.48 a | ||
고 찰
개량머루의 액아는 기존에 보고된 다른 품종과 형태와 발달과정은 유사하였으나 괴사가 거의 발생하지 않아 액 아의 발달이 매우 우수하였다. Choi 등(2007)은 캠벨얼 리의 주아괴사율이 28.6~96.8%이며 Kim 등(2011)은 홍 이슬의 정상눈은 8%에 불과하다고 보고했지만, 개량머 루의 액아는 괴사가 발생하지 않아 다른 품종에 비해 눈의 소질이 우수하고 충실하였다.
개량머루의 액아는 신초의 등숙이 완료되는 10월까지 크기가 계속 증가했다. 반면에 Kim 등(2011)의 보고에 의하면 홍이슬의 액아는 관찰시기에 따라 크기가 작아진 다고 보고하여 정반대의 결과를 보였다. 이러한 결과는 홍이슬의 액아에 괴사가 심하게 발생하여 시간이 경과함 에 따라 눈의 크기가 작아지지만 개량머루는 액아의 괴 사가 전혀 발생하지 않았기 때문에 시간이 경과할수록 눈의 크기가 증가하였다. 포도에서 기부에 위치한 1번 눈은 크기가 작고 휴면이 심한 잠아로 괴사발생이 심하 지만(Keller, 2010), 개량머루의 1번 눈은 다른 위치의 눈과 크기 차이가 없고 괴사도 발생하지 않아 충실했다.
눈의 충실함은 이듬해 신초 및 꽃송이 확보와 밀접한 관계를 보여 캠벨얼리와 홍이슬의 경우 액아 괴사로 인 해 신초 및 꽃송이 확보가 어려워 생산량의 확보가 어 렵다(Choi 등, 2007; Kim 등, 2011). 액아의 괴사 발생 은 수체내 양분의 영향을 받아 다른 기관과의 양분경합 에 의해 발생한다(Vasudevan 등, 1998). 하지만 개량머 루는 강한 수세에도 불구하고 액아의 괴사가 전혀 발생 하지 않고, 특히 위치에 관계없이 소질이 매우 우수하여 수형을 구성함에 있어 장초 혹은 단초 전정 모두 신초 와 꽃송이 확보가 가능할 것으로 판단되었다.
안성에서 재배된 개량머루의 과실은 변색 1주전인 만 개 후 9주를 기점으로 당의 축적과 산의 감소가 시작되 었다. 일반적으로 성숙기 포도는 과당과 포도당으로 이 루어져 있고 일부 품종에서는 자당이 축적된다고 보고되 었다(Lee 등, 2003; Matsumoto 등, 2007; Matsui 등, 1986). 축적되는 당에 따라 환원당 축적형과 비환원당 축적형으로 구분할 수 있으며, 개량머루는 과당과 포도 당이 축적되므로 환원당 축적형 품종으로 구분할 수 있 었다(Shiraishi, 2000). 당이 축적됨에 있어 포도당과 과 당은 동일한 패턴으로 증가하였고 포도당과 과당의 비율 이 1에 가까워져 Shiraishi(2000)가 보고한 거봉의 당 조 성과 가장 유사하였다. 개량머루는 착색이 우수하여 당 도와 산도가 성숙이 완료되기 전 착색이 완료되는 특성 으로 인해 착색 정도를 기준으로 수확하는 경우 성숙이 완료되지 못한 과실이 수확될 가능성이 높다. 일부 포도 품종에서는 성숙한 과실은 포도당과 과당의 비율이 1에 가까워지므로 수확기준으로 활용하고 있으며 이를 이용 한 수확기준 설정이 개량머루에서 적합할 것으로 판단되 었다(Amerine과 Thoukis, 1958).
Lee 등(2003)과 Matsui 등(1986)은 포도는 사과산이 가장 많다고 보고하였으며, 개량머루 또한 사과산의 함 량이 가장 많아 Soyer 등(2003)과 Siraishi(2000)가 보고 한 유기산의 조성과 일치하였다. 개량머루의 성숙이 진 행될수록 적정산도는 사과산에 직접적인 영향을 받아 산 도가 감소되는 시기적 변화가 일치하였다.
수정직후에 배주의 퇴화가 관찰되었고, 수정이 완료된 개화 1주 후에도 계속해서 퇴화되는 배주가 관찰되어 개량머루에서 배주의 퇴화는 수정전과 후에 모두 진행되 는 것으로 생각되었다. 포도는 품종 특성에 따라 성숙기 종자의 수에 차이가 나며 이와 더불어 동일한 품종에서 하나의 과립에서도 종자의 크기에 차이를 보이는 경우가 있어 이는 배주가 퇴화하는 시기에 따라 달라지는 것으 로 생각된다.
성숙기에는 개화기에 비하여 외표피에서 유관속조직까 지의 세포층 수가 약 2.1배 증가하였고 두께가 3.8배 증 가하여 세포분열과 세포비대가 이루어진 것을 확인하였 다. 또한 과실비대 초기에는 세포비대와 세포수의 증가 가 동시에 이루어진 것이 관찰되었으며 만개 후 2주부 터 성숙기까지는 세포층 수의 변화가 없어 세포분열은 만개 후 14일 전후에 중단되었고 세포비대는 계속 진행 되었다. Hardie 등(1996)는 Sauvignon Blanc 포도의 세 포분열 정지기는 28일 전후라고 보고하여 개량머루의 세포분열기는 다른 품종에 비해 매우 짧은 것이 확인되 었다. 성숙기의 과피는 한 층의 epidermis 세포와 7~10 층의 hypodermis 세포로 구성되었고, 과피 세포의 타닌 이 다른 포도 품종에 비해 적은 것으로 관찰되었다(Lee 등, 2010). 기존 연구에서 과실 비대 중에 epidermis 세 포의 타닌이 완전히 사라졌다가 재형성 되는 경우는 보 고되었으나(Park과 Park, 2000) 이와 같이 hypodermis 층을 구성하는 세포에서 타닌이 완전 분해된 후 재합성 되는 것은 다른 과실에서는 보고된 바가 없어 이에 관 한 연구가 향후 이루어져야 할 것으로 생각되었다.
