

서 론
낙엽과수의 휴면은 식물체 내부적으로는 생리적 변화 가 계속 일어나고 있으나 외관상 생장이 멈춘 상태를 말한다(Saure, 1985). 휴면은 유기 요인에 따라 외재휴면 (para-dormancy), 내재휴면(endo-dormancy), 환경휴면 (eco-dormancy) 단계로 구분하고 있으며, 외재휴면은 눈 이 식물체의 다른 부위와의 상관적 억제작용 때문에 발 아하지 못하는 상태(Lang 등, 1987)로써, 정단신초를 적 심하거나 적엽처리 등에 의해 액아나 정아의 재발아가 일어날 수 있다.
과거 인위적 적엽처리는 사과의 과실 성숙기간 중 과 실의 착색 증진과 품질 향상을 위한 목적으로 연구되어 왔으나(Yim과 Lee, 1999; Choi 등 2000), 적엽 시기가 너무 빠르거나 정도가 심하게 되면 과실의 당 함량과 비대량 감소 등 부정적인 영향이 나타나게 된다(Sang 등, 2005).
최근 우리나라에서는 지구온난화에 의한 기상재해가 빈번히 발생하면서 강한 바람과 태풍에 의한 조기낙엽 피해가 증가하고 있다. 과수의 생육시간 중 잎 손실은 당해 연도에 과실크기, 색도, 당도를 감소시켜 과실의 품질을 저하시키며(Choi 등, 2002), 화아분화, 수체 저장 양분 및 꽃눈 발달에 나쁜 영향을 미친다(Jeong 등, 2005). 또한, 조기낙엽에 의해 수체의 저장양분이 감소하 여 이듬해 개화와 결실이 감소하게 되며(Kang과 Ko, 1976), 익년 내한성과 화아분화에도 영향을 주는데 낙엽 율이 높을수록 가용성 양분저장이 감소하여 내한성과 개 화율이 감소한다(Sagong 등, 2011).
본 연구에서는 사과나무의 생육기간 중 태풍과 우박 등 극한기상 조건에 의한 조기낙엽 피해를 가정하여 인 위적으로 적엽을 하였으며, 적엽처리 직후 2주(14일) 동 안 남아있는 잎에서 광합성 능력과 탄수화물을 조사하여 조기낙엽 발생 초기의 수체 내에서 일어나는 저장양분의 변화를 알아보고자 수행하였다.
재료 및 방법
1. 시험재료 및 적엽처리
시험재료는 2016년 3월 15일 전라북도 완주군에 소재 한 농촌진흥청 국립원예특작과학원에서 3년생 사과‘후지’ (Malus domestica Borkh.)/M9을 포트에 재식하였다. 재 식방법은 식토, 육묘용 상토, 퇴비를 5:3:2 비율로 혼합 한 토양을 50L 포트에 채우고 각 포트마다 묘목을 1주 씩 심었다. 시험에 사용한 사과나무는 착과시키지 않았 다. 적엽 수준은 80%, 50%, 0%(대조구)로 8월 2일에 수행하였고, 완전임의배치 3반복으로 시험구를 배치하였 다. 적엽처리는 대조구의 전체 엽면적을 고려하여 수관 전체에서 골고루 비율에 맞게 전정가위를 이용하여 제거 하였다. 탄수화물 분석을 위한 시료채취는 적엽처리 후 2일, 7일, 10일, 14일에 각각 실시하였으며, 뿌리의 저장 양분 분석을 위한 시료는 적엽처리 30일 후 채취하였다. 조사된 데이터는 통계분석 패키지인 SAS 9.2(SAS Institute, Inc., Cary, USA)를 이용하여 분석하였다.
2. 수체 생육, 재발아율 및 탄수화물 변화
수체 생육 조사는 적엽처리 30일이 경과한 후 접목부 위로부터 10cm 윗부분에서 주간부 직경과 수폭을 측정 하였으며, 수고는 지면을 기준으로 측정하였다. 눈의 2 차 재발아율은 각 시험수의 전체 눈 수를 조사한 후 발 아한 눈 수를 백분율로 계산하였다.
탄수화물 분석은 분쇄한 시료 0.5g을 500mL 삼각플라 스크에 넣고 0.7N HCL 20mL을 첨가한 후, 100°C의 항온수조에서 2시간 30분 동안 중탕한 후 용액을 여과 지로 여과한 다음 증류수로 전체 용량이 100mL이 되도 록 하였다. 분해액 3mL에 dinitrosalicylic acid 5mL을 첨가한 후 10분간 끓인 후 증류수로 전체 용량을 50mL 로 하여 발색시킨 후 흡광분광분석기(UV0250 1PC, Shimadzu, Japan)를 이용하여 550nm에서 흡광도를 측정 하였다.
유리당 분석은 시료 0.5g에 3차 증류수 25mL를 첨가 하여 100°C에서 30분간 추출한 후 1분간 균질화시켰다. 4°C에서 12,000rpm으로 20분간 원심분리하여 얻어진 상 등액을 C18 cartridges (Sep-Pak, Waters Co., USA)로 여과하여 분석하였다. HPLC (M717, Waters Co., USA) 분석에 사용한 칼럼은 sugarpak TM (6.5×300mm, Waters Co., USA)으로, 이동상은 HPLC용 증류수이고, 이 동속도는 0.5mL/min이였다. 유리당의 검출은 refractive index detector (Waters Associates Differential Refractometer R410, Waters Co., USA)로 하였으며, 유리당 함량은 시 료 중의 각 유리당과 동일한 표준물질(Sigma Chemical Co. Ltd., USA)을 이용하여 검량선을 작성하였다.
결과 및 고찰
1. 수체 생육 및 재발아율
생육기간 중 조기낙엽이 되었을 때를 가정하여 사과 ‘후지’/M9 품종을 인위적으로 적엽처리한 후 수체 생리 반응을 관찰하였다. 적엽처리 30일 후 수체 생장량(수고, 수폭, 간경)의 변화에는 유의한 차이가 없었다. 반면 눈 의 2차 재발아율은 적엽처리 후 80% 처리구에서는 82.7% 로 매우 높게 나타났으며, 50%처리구, 무처리구는 각각 45.9%, 2.1%로 매우 유의한 차이를 나타냈다(Table 1).
사과의 꽃눈 원기는 여름에 형성되며 가을에 눈이 정 상적으로 휴면에 진입하여야 정상적인 눈으로 발달하게 된다(Rhee와 Ko, 1973). 이러한 사과의 휴면은 ‘시나노 스위트’에서 9월 중순 이후에 이루어지며(Han 등, 2016), ‘후지’와 ‘홍로’는 9월 말, ‘쓰가루’는 9월 초에 내재휴면에 진입하는 것(Kweon 등, 2013)으로 알려져 있다. 그러나 본 연구에서는 이보다 빠른 외재휴면 상태 에서 인위적으로 조기낙엽 되어 눈이 정상적으로 내재휴 면으로 진입하지 못하였으며, 이로 인하여 80% 처리구 에서 생육기간 중 눈의 2차 재발아가 높게 유도된 것으 로 판단된다. 눈의 2차 재발아는 휴면과 관련이 있으며 정상적으로 내재휴면이 유도될 경우 눈의 2차 생장이 일어나지 않게 된다. 이러한 적엽처리에 의해서 눈의 2 차 재발아가 일어나고, 동화양분이 소모되어 이듬해 꽃 눈의 감소 및 저장양분 부족으로 내한성이 떨어진다 (Kang과 Ko, 1976).
Table 1. Growth increment of plant height, plant width, stem diameter and regrowth of buds in ‘Fuji’/M9 apple trees at 30 days after defoliation treatments.
| Defoliation | Plant height (cm) | Plant width (cm) | Stem diameter (mm) | Regrowth of buds (%) |
|---|---|---|---|---|
| 0% | 11.3 aZ | 1.7 a | 3.8 a | 2.1 c |
| 50% | 18.3 a | 8.3 a | 5.8 a | 45.9 b |
| 80% | 15.7 a | 10.0 a | 5.6 a | 82.7 a |
Fig. 1은 적엽처리 후 정단아의 재발아한 모습으로 무 처리구에서는 거의 재발아가 일어나지 않았으나, 50% 처리구에서는 재발아하여 잎이 전개되었으나 80% 처리 구보다 늦게 일어나는 양상이었다. 적엽처리 정도가 심 할수록 재발아가 급격히 일어난 것은 재발아 된 신초 잎들이 수체 생장에 필요한 동화산물의 축적을 위하여 source 잎으로써 역할을 하는 것으로 판단된다.

2. 탄수화물 변화
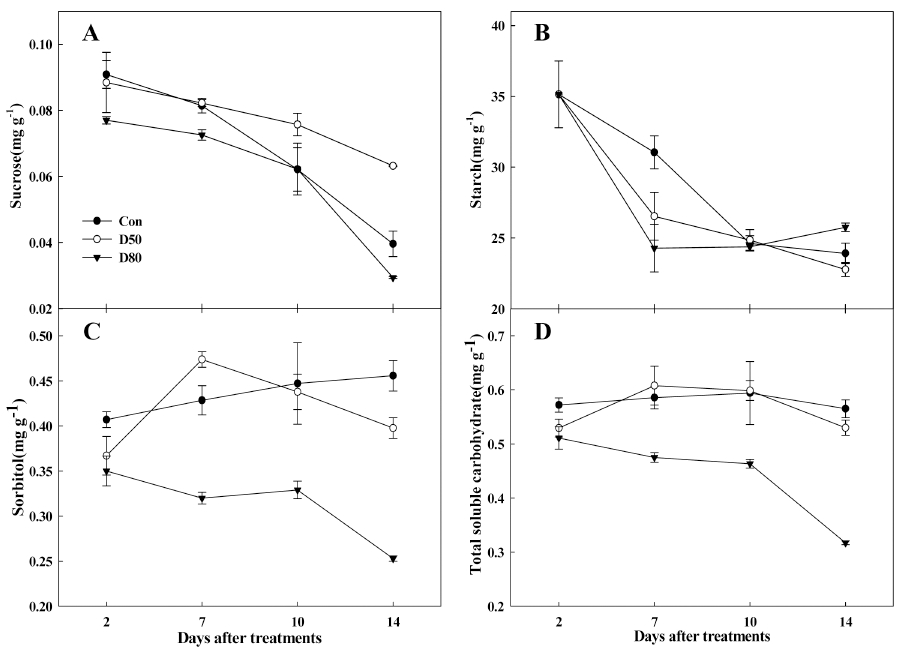
일반적으로 사과나무를 포함한 Rosaceae과 과종은 광 합성 초기에 동화산물로 sorbitol을 생성하며, 잎의 유리 당은 대부분 sorbitol이 차지하고 있다(Loescher 등, 1982). 이러한 유리당과 전분 함량의 변화를 적엽처리 후 14일 동안 조사하였다(Fig. 2). 대조구와 50% 처리구 의 총 유리당은 0.5mg·g-1부터 0.6mg·g-1사이에서 일정하 게 유지되었다. 반면, 80% 처리구에서는 2주동안 꾸준 히 0.5mg·g-1에서 0.3mg·g-1로 약 40% 감소하는 경향이었 으며(Fig. 2D) 이는 sorbitol의 함량이 0.3mg·g-1에서 0.2mg·g-1로 줄어드는 것과 같은 양상을 나타냈다(Fig. 2C). 80% 처리구에서 sucrose의 함량은 유의한 차이 없 이 일정하게 감소하였으며(Fig. 2A) 전분의 함량은 7일 까지 줄어든 후 일정하게 유지되었다(Fig. 2B). Faust 등 (1968)은 세포분열이 왕성하거나 대사작용이 활발한 조 직이나 기관으로 양분의 이동이 많아진다고 하였으며, 본 연구 결과와 같이 적엽처리 후 source 잎의 유리당이 감소하는 것으로 판단하였을 때 눈의 2차 재발아에 의 해 source 잎으로부터 sink 눈으로 양분이 이동한 것으로 추정하였다. 또한 적엽처리로 인하여 sink-source가 조절 되었을 경우 잎에서 전분과 비구조 탄수화물의 함량이 감소하여 삼투포텐셜이 증가할 것(Rui와 Quebedeaux, 2003)이라 보고하였다. 2차 재발아한 sink 눈으로 양분 이 이동함에 따라 잎조직 내에서 삼투포텐셜이 증가하게 되고 수분포텐셜이 낮아질 것으로 추정된다.
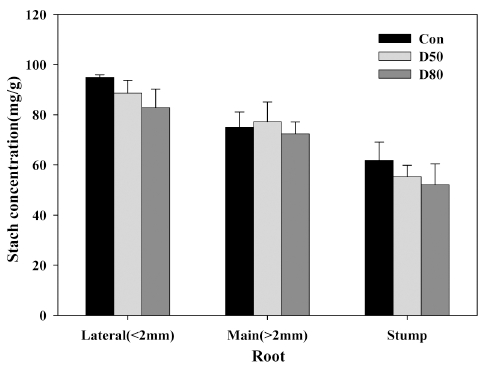
인위적 적엽을 통해 생육기 조기낙엽이 발생했을 경우 를 가정하여 뿌리의 저장양분 중 전분함량을 측정하였다 (Fig. 3). 적엽처리 정도가 심할 수록 뿌리의 전분함량은 감소하는 경향을 나타냈으나 통계적으로 유의한 차이는 없었다. 낙엽과수의 경우 신초의 생장이 멈춘 시점부터 저장양분의 축적이 뿌리와 같은 주요 저장기관으로 이루 어진다(Oliveira와 Priestley, 1988). 특히 탄수화물의 저 장형태로서 전분의 축적은 이듬해 생장을 위해 뿌리, 줄 기와 같은 영구조직에 축적된다. 그러나 생육기 조기낙 엽은 저장 탄수화물의 축적을 불량하게 하고 저장 기관 중 뿌리가 가장 민감하게 반응한다(Loescher 등, 1990) 고 하였다. Cruz-castillo 등(2010)은 적엽처리에 의한 비 구조 탄수화물과 전분 함량을 수체의 각 부위별로 조사 한 결과 1년생 가지와 주간부에서는 유의하게 감소하였 으나, 잔뿌리에서는 감소하는 경향은 있었으나 유의한 차이는 찾을 수 없었다고 보고와 유사하였다. 본 연구는 적엽처리 30일 후 뿌리의 탄수화물 및 전분 함량 변화 를 측정한 것으로 뿌리의 저장양분 감소와 수체 약화의 상관관계를 좀더 명확하게 해석하기 위해서는 향후 내한 성 평가 등의 추가 연구가 필요할 것으로 판단된다.
3. 광합성 변화
적엽처리에 따른 광합성 변화를 14일 동안 측정한 결 과는 Fig. 4와 같다. 광합성 속도(Pn)는 대조구에서는 11.9μmol·m-2·s-1에서 5.2μmol·m-2·s-1으로 감소하였으나 80% 처리구에서 측정기간 동안 11.1μmol·m-2·s-1에서 14.3μmol·m-2·s-1으로 꾸준히 증가하였으며 50% 처리구 에서는 처리 후 7일까지 증가하다 감소하는 경향이었다 (Fig. 4A). 기공전도도는 모든 처리구에서 7일까지 증가 하는 양상이었으나, 7일 이후에도 80% 처리구는 0.31mol·m-2·s-1까지 꾸준히 증가하는 반면 대조구, 50% 처리구에서는 감소하였다(Fig. 4B). 일반적으로 광합성속 도와 기공전도도는 정의 상관관계를 나타내고 있으며 기 공전도도는 잎의 삼투포텐셜이 증가할 경우 기공이 열리 게 되어 증가하는 것으로 알려져 있다. 또한, 적엽처리 에 의해 sink-source 조절에 따른 광합성률의 증가는 잎 조직의 삼투포텐셜 증가와 연관이 있다고 알려져 있다 (Rui와 Quebedeaux, 2003).
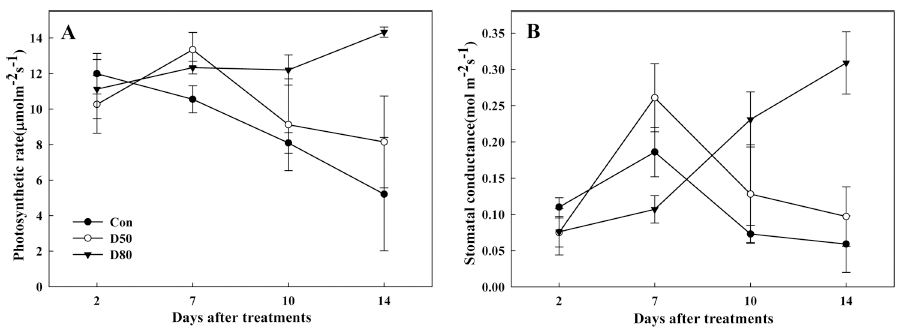
눈의 재발아에 의하여 성숙한 엽에서의 유리당과 전분 의 감소가 삼투압을 증가시키고, 증가된 삼투압에 의해 서 기공이 열리게 되어 광합성이 증가한 것으로 판단된 다. Priestley (1987)에 의하면 광합성 산물의 이동은 광 합성 산물의 수요기간인 sink activity, sink size가 관계 되어 있으며, 특히 sink activity에 의해 좌우된다고 하였 다. 결과적으로 조기낙엽에 의한 피해는 휴면에 들어가지 못한 눈의 2차 재발아에 따라 탄수화물 요구가 증가함으 로써 양분의 소모가 일어나게 되는 것으로 판단된다. 따 라서 이러한 조기낙엽 피해를 경감시키기 위해서는 눈의 재발아와 같은 불규칙적인 생장을 억제시킴으로써 해결 할 수 있을 것으로 예상되며, 향후 이상발아에 대한 억 제 방법 연구가 이루어져야 할 것으로 판단된다.
