

서 론
시설 내 작물의 연중 생산에 따른 묘에 대한 수요 증 가, 농촌인구 감소와 노령화 등 작물 생산환경의 변화에 따라, 최근 작물의 생산, 특히 채소 생산에 있어서는 육 묘와 재배의 분업화 및 전문화가 이루어지고 있다. 재배 농가는 직접 묘를 키우기 보다는 전문 공정육묘장에서 생산된 묘를 구입하여 이용하는 비중이 지속적으로 증가 하고 있다. 2014년 재배농가의 구입묘 이용현황 조사에 의하면, 채소묘의 구입 비중은 토마토 74%, 수박 72%, 오이 65%, 가지 63%, 고추 56%로, 과채류의 묘 구입 비중이 높았다(Park 등, 2014).
재배농가는 주로 묘의 품질이 우수한 공정육묘장을 선 택하기 때문에, 인근의 공정육묘장뿐만 아니라 다른 지 역의 공정육묘장에서 묘를 구입하기도 한다(Park 등, 2011). 따라서, 묘의 운송은 공정육묘장이 위치한 지역 내에서뿐만 아니라 전국적으로 이루어지고 있다. 공정육 묘장에서 생산된 묘는 일반적으로 플러그 트레이에 심겨 진 채 종이상자에 포장되어, 탑차나 트럭 등에 탑재되어 상온에서 운송되고 있다.
공정육묘장 시설 내 적절한 환경조건에서 육묘된 묘는 재배농가로의 운송과정에서 급격한 환경변화 또는 저온 이나 고온 조건에 노출되기 쉽다. 이러한 운송시의 환경 조건은 묘의 품질에 영향을 미쳐, 정식 후 생육 및 수량 등에 영향을 준다. 특히 운송과정에서 상온의 포장된 상 자 내부와 같이 암조건 하에서는 묘의 품질이 빠르게 저 하된다(Kubota와 Kroggel, 2006; Kubota 등, 2004). Kubota와 Kroggel(2006)에 의하면, 19°C의 상온 조건에 서 4일간 운송된 토마토묘는 정식 후 꽃이 퇴화되거나 착과가 정상적으로 이루어지지 않고 과실 발육이 늦어져 수량이 낮아졌다. 반면, 저온(6-13°C) 및 약광 조사 (12μmol·m-2·s-1PPF) 조건에서 운송하였을 때는 외관상의 품질이 유지되었으며 이후 높은 광합성 능력을 유지하였 으며 수확시기의 지연이나 수량감소 등의 문제가 없었다.
구입묘 이용시 정식 후 안정적인 생육 및 수량확보를 위해서는, 고품질의 묘 생산과 함께 유통환경에도 주목할 필요가 있다. 따라서 본 연구는 토마토 묘의 운송 기간 중 품질유지를 위한 온도 조건을 구명하고자 수행되었다.
재료 및 방법
실험재료 및 육묘
실험재료로 토마토(Solanum lycopersicum) 접목묘를 이용하였다. 접수품종은 ‘슈퍼도태랑((주)코레곤, 한국)’, 대목품종은 ‘B블로킹(한국다끼이㈜, 한국)’이었다. 원예 용 상토(BM2, Berger Group Ltd., 캐나다)를 충진한 40 공 플러그 트레이((주)범농, 한국)에 토마토 종자를 파종 하고 충분한 양의 물을 두상관수 하였다. 발아를 촉진시 키기 위해 플러그 트레이 윗면에 비닐랩을 씌운 후, 온 도 27°C로 설정된 발아실에 4일간 두었다. 이후 유리온 실 내 육묘베드 위로 옮겨 육묘하였다. 본엽 4매 정도가 전개되었을 때 합접 방법으로 접목한 후, 온실 내 육묘베 드 위에 설치된 활착상 내에서 일주일 동안 활착시켰다. 접목 활착이 끝난 후에는 일반 육묘베드에서 육묘하였다. 활착기간을 제외한 육묘기간 중 토마토의 생육과 날씨에 따라 물과 육묘용 양액(전기전도도(EC) 1.4dS·m-1, ‘한방’ 육묘용 양액, N-P-K-Ca-Mg = 8.0-2.4-2.4-4.8-1.6me·L-1, (주)코씰, 한국)을 두상관수 하였다.
정식 전 온도 처리
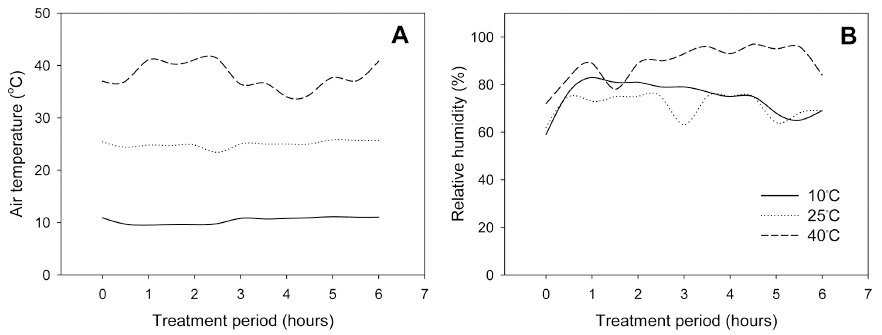
토마토묘의 운송시 온도 조건이 정식 후 생육 및 수량 에 미치는 영향을 검토하기 위해, 본엽이 9-10매 정도 전개하고 1화방의 1번화가 피기 직전 또는 핀 토마토 접목묘를 이용하여, 온도 처리수준 및 시간을 달리하여 처리하였다. 온도처리는 온도조절이 가능한 식물생장상 (HB-301LP, 한백과학, 한국)을 이용하였다. 3대의 식물 생장상의 온도를 각각 10, 25, 40°C로 설정하였다. 오전 10시에 토마토묘를 온실에서 암조건의 식물생장상으로 옮겨 2, 4, 6시간 처리하였다. 처리기간 중 기온과 상대 습도는 Fig. 1과 같았다. 온도 10와 25°C 처리구의 경우 상대습도가 60-80% 범위였으나, 40°C 처리구의 경우 100%에 가까웠다. 온도 처리 종료 후 토마토묘를 일정 기간 차광조건에서 순화하였다. 처리기간 중 온실에서 육묘된 토마토묘를 대조구로 하였다.
정식 및 재배
온도 처리 종료 후 식물생장상에서 꺼내어 관리동 그 늘진 곳에 두었던 토마토묘를 다음날 아침 비닐하우스에 정식하였다. 비닐하우스 내에 세 줄의 이랑을 만들고, 정식 전에 각 이랑에 두 줄의 점적테이프를 설치, 검정 색 필름으로 멀칭하였다. 각 이랑에 토마토묘를 40cm 간격으로 2줄로 심어 재배하였다. 시험구는 난괴법 3반 복으로 배치하였으며, 각 반복마다 처리별로 6주의 식물 체를 심었다. 토마토는 측지를 제거하여 원줄기만 재배 하였다. 한 화방의 과실수를 4개 이하로 조절하였고, 2 화방까지 수확하였다.
조사 및 통계분석
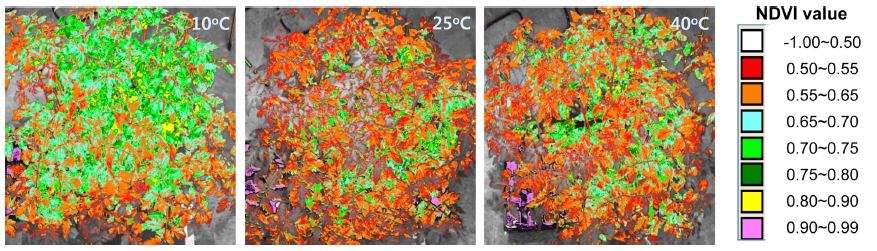
온도처리 종료 후 근적외선 디지털 카메라(D100 IR, Nikon, 일본)를 이용하여 처리별 토마토묘를 촬영, 정규 화 식생지수(Normalized Difference Vegetation Index, NDVI)를 구하였다(Govaerts와 Verhulst, 2010).
처리 종료 직후 처리당 10주씩의 식물체를 채취하여, 지상부와 지하부의 생체중을 측정하였다. 조사 후 시료 를 80°C로 설정된 열풍건조기(DS-89, 다솔과학, 한국)에 서 3일이상 건조하여 건물중을 측정하였다. 정식 후 초 기생육 검토를 위해, 정식 17일 후 반복마다 처리별로 2 주씩의 식물체를 채취하여, 초장, 경경, 엽수(황화엽 및 낙엽된 잎 포함), 엽면적, 생체중, 건물중을 조사하였다. 반복마다 처리별로 3주의 식물체에 대해 토마토 1화방 의 착화절위 및 화수를 조사하였으며, 정식 53일 후 1화 방의 수량을 조사하였다.
수집된 자료는 시그마플롯(v.11.0, Systat Software Inc., UK)과 SAS 통계프로그램(v.9.2, SAS Institute, USA)을 이용하여 분석하였다.
결과 및 고찰
온도처리 종료 후 근적외선 디지털 카메라(D100 IR, Nikon, 일본)로 촬영하여 분석한 토마토묘의 NDVI 영상 은 Fig. 2와 같다. 식물은 건강상태, 활력에 따라 가시광 선 영역과 근적외선 영역의 파장대에서 특징적인 분광특 성을 나타낸다. 이러한 특성을 활용하여 NDVI 값을 구 할 수 있는데, NDVI 값은 -1에서 1사이의 범위의 값을 가지며, 값이 클수록 활력이 좋고 건강한 식물이며, 반 대로 스트레스를 받은 경우엔 상대적으로 낮은 값을 갖 는다(Govaerts와 Verhulst, 2010). NDVI 값은 광합성, 수량 등과 정의 상관을 갖는 것으로 보고되고 있다(Ma 등, 2001). 10°C 처리구의 토마토묘는 전반적으로 NDVI 값 0.7-0.8범위의 연두색에서 초록색을 띤 반면, 25, 40°C 처리구의 토마토묘는 NDVI 값 0.5-0.65 범위 의 주황색에서 붉은 색을 띠어, 10°C 처리구에 비해 25, 40°C 처리구에서 더 낮은 NDVI 값을 나타냈다.
처리 종료 직후 토마토묘의 지상부 생체중과 지상부, 지하부 건물중은 처리간에 차이가 없었지만, 지하부 생 체중은 처리시간에 상관없이 온도 조건의 영향을 받아, 40°C 처리구에서 가장 낮은 값을 보였다(Table 1). Shishido와 Kumakura(1994)에 의하면, 근권 온도 상승에 의해 토마토의 증산량 및 호흡량이 증가하며, 10°C에서 22°C로 온도 상승시 토마토의 근권부 호흡량은 3배가 증가하였다. 따라서 40°C 처리구에서 지하부 생체중 감 소는 암상태의 고온조건에서의 증산량 및 근권부 호흡량 증가에 기인한 것으로 추정된다.
Table 1.
The fresh and dry weight of tomato transplants as affected by temperature treatment fo simulte transportation.
| Treatment | Fresh weight (g) | Dry weight (g) | |||
|---|---|---|---|---|---|
| Temperature (°C) | Period (hours) | Shoot | Root | Shoot | Root |
| Control | 13.36abz | 1.27ab | 1.18a | 0.09a | |
| 10 | 2 | 12.56a-c | 1.40ab | 1.03a | 0.10a |
| 4 | 12.37a-c | 1.26a-c | 1.05a | 0.09a | |
| 6 | 13.08a-c | 0.83cd | 1.22a | 0.13a | |
| 25 | 2 | 13.83a | 1.61a | 1.15a | 0.13a |
| 4 | 13.47a | 1.30ab | 1.09a | 0.11a | |
| 6 | 11.31c | 1.45ab | 1.08a | 0.10a | |
| 40 | 2 | 12.08a-c | 1.00b-d | 1.04a | 0.13a |
| 4 | 12.17a-c | 0.78d | 1.03a | 0.14a | |
| 6 | 11.44bc | 1.01b-d | 1.02a | 0.13a | |
| P value | |||||
| Temperature (A) | 0.1334 | 0.0001 | 0.3278 | 0.1526 | |
| Period (B) | 0.1842 | 0.0657 | 0.6326 | 0.9369 | |
| A × B | 0.1124 | 0.1226 | 0.3230 | 0.1758 | |
정식 5-7일 후에는 40°C 처리구 토마토묘의 아래잎(첫 번째-세번째 본엽 사이)에서 황백화 및 잎마름 증상이 발생하였다(Fig. 3). Kubota와 Kroggel(2006)는 육묘장에 서 재배농가로의 토마토묘 운송시 4일간의 운송기간 중 암조건 또는 19°C 온도조건에 놓인 토마토묘를 정식하 였을 때, 정식 일주일 후 잎에서 황백화 및 괴사 증상이 나타났다고 보고하였다. 그들은 이러한 증상의 발생이 4 일간의 암조건에서 밝고 따뜻한 온실로의 환경변화에 의 한 광저해(photoinhibion)과 관련이 있을 것으로 추정하 였다. 본 실험에서는 모든 처리를 동일하게 암조건으로 처리하였음에도 불구하고, 40°C 처리구에서만 황백화 및 잎마름 증상이 발생하였다. 처리가 끝난 후에는 관리동 그늘진 곳에 두었다가 다음날 정식을 하였기 때문에, 갑 작스러운 광환경 변화에 의한 증상이기 보다는 처리기간 중 고온이 증상의 발현에 영향을 준 것으로 추정된다.

정식 17일 후 생육 조사 결과, 초장, 엽수, 지상부 생 체중, 건물중에 있어서는 처리가 차이가 없었으나, 엽면 적은 처리 온도가 높을수록 감소하는 보여, 40°C 처리구 에서 가장 낮은 값을 보였다(Table 2). 이러한 결과는 황 화 및 괴사에 의한 하엽의 낙엽에 기인한 것으로 추정 된다.
Table 2.
The growth of tomato transplats as arrected by temperature treatment to simulate transportation at 17 dats after transplanting.
| Treatment | Shoot length (cm) | Number of leaves | Leaf area (cm2) | Shoot weight (g) | ||
|---|---|---|---|---|---|---|
| Temperature (°C) | Period (hours) | Fresh | Dry | |||
| Control | 82.6az | 16.3a | 1,155b | 93.1a | 12.0a | |
| 10 | 2 | 77.9a | 15.0a | 1,152b | 89.8a | 11.1a |
| 4 | 78.2a | 15.3a | 1,194ab | 96.0a | 11.2a | |
| 6 | 79.6a | 17.0a | 1,333a | 109.1a | 12.1a | |
| 25 | 2 | 78.6a | 15.8a | 1,192ab | 92.4a | 11.4a |
| 4 | 79.2a | 16.0a | 1,173ab | 89.2a | 10.7a | |
| 6 | 74.9a | 16.0a | 1,060b | 86.7a | 10.5a | |
| 40 | 2 | 76.1a | 16.0a | 1,112b | 87.1a | 10.1a |
| 4 | 77.2a | 15.8a | 1,067b | 84.1a | 10.0a | |
| 6 | 78.5a | 15.8a | 1,114b | 94.4a | 11.0a | |
| P value | ||||||
| Temperature (A) | 0.6352 | 0.9084 | 0.0340 | 0.0797 | 0.0909 | |
| Period (B) | 0.8939 | 0.1949 | 0.8774 | 0.2329 | 0.5466 | |
| A × B | 0.3309 | 0.1593 | 0.1065 | 0.2763 | 0.5488 | |
운송 온도조건에 따른 토마토 묘의 1화방의 착화 절위 및 착화 개수를 조사한 결과, 1화방 착화 절위는 처리에 따른 영향을 받지 않았다. 토마토의 화아 분화는 환경조 건과 식물체의 발육상태에 따라 다르지만, 일반적으로 본엽이 8매 정도 전개되기까지의 육묘기간 중에 이미 3 화방까지의 화아 분화가 이루어진다(RDA, 2014). 따라 서 1화방의 착화 절위는 운송 중 온도처리 조건에 상관 없이 10절 내외에 위치하였다. 그러나 1화방의 착과 개 수는 운송 중 온도처리 조건의 영향을 받아, 다른 처리 구가 착과수가 3개 내외였던 반면, 40°C 6시간 처리구 의 경우 착과수가 감소하여 착과수가 2개였다(Fig. 4).
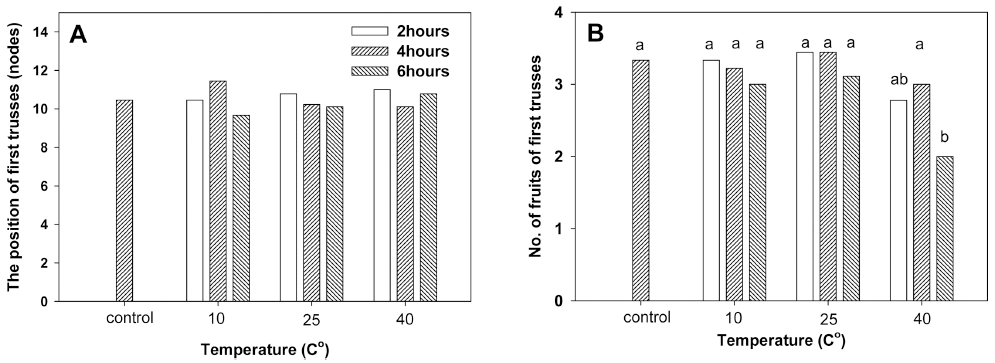
토마토의 생육적온은 주간 24-26°C, 야간 18-20°C이다. 토마토의 생식생장 기간 중 주간 30°C, 야간 21°C의 고 온, 또는 15°C 이하의 저온에 의해 생식생장이 저해되며, 주간온도보다 야간온도에 대해 더 민감하게 반응한다 (Osorio 등, 2014). 특히 토마토의 화기는 고온 스트레스 에 매우 민감하여 30°C 이상의 고온 조건, 특히 40°C 이상에서는 몇 시간 동안의 노출로도 화분낭이 기형화되 고 화분의 활력은 저하하여, 정상적인 착과가 이루어지 지 못해 착과율이 낮아진다(De Storme과 Geelen, 2014; Giorno 등 2013; Paupière 등, 2014; Pressman 등, 2002). 2-8mm길이의 토마토 꽃눈에서는 화분모세포의 감수분열을 통해 소포자 형성과 화분형성이 순차적으로 이루어지는데, 개화 8-13일 전에는 고온에 특히 민감하 여 이 시기에 고온에 노출되면 전분 및 당의 감소에 따 른 화분 발달의 장애로 착과가 크게 감소하게 된다 (Giorno 등 2013; Pressman 등, 2002). 일반적으로 국내 에서 토마토묘가 육묘장에서 재배농가로 운송되는 시기 의 토마토의 묘령은 첫번째 화방의 1번화가 피기 전후 단계로, 이 시기에는 고온에 의해 화기가 영향을 받기 쉬워 착과 감소로 이어질 우려가 있다. 본 실험에서도 6 시간의 고온 처리에도 착과가 영향을 받아 착과수가 감 소하였다.
토마토 화방 내 과실의 발달은 기부쪽 1번과부터 선단 부쪽으로 이루어져, 1번과가 가장 크고 선단부로 갈수록 과실이 작다(Kubota와 Kroggel, 2006). 본 실험에서는 운송 온도 및 처리 시간이 과실 무게에 영향을 주어, 1 번과와 3번과의 경우 40°C 처리구와 25°C 6시간 처리구 의 무게가 다른 처리구에 비해 작았으며, 이러한 무게 감소는 1화방의 수량감소로 이어졌다(Table 3). 40°C 6 시간 처리구의 1화방 수량은 대조구의 40% 정도였다. 2 화방의 수량은 처리간에 차이가 없었다.
Table 3.
The development and yield of the first trusses as affected by temperature treatment to simulate transportation.
| Treatment | Fruit size (g) | Yield of the first trusses (g) | ||||
|---|---|---|---|---|---|---|
| Temperature (°C) | Period (hours) | First | Second | Third | Fourth | |
| Control | 156abz | 162a | 217a | 178a | 587a | |
| 10 | 2 | 146a-c | 148a | 152bc | 108a | 478a |
| 4 | 153a-c | 175a | 186ab | 166a | 547a | |
| 6 | 177a | 164a | 218a | 191a | 556a | |
| 25 | 2 | 146a-c | 179a | 168a-c | 189a | 579a |
| 4 | 135a-c | 152a | 161a-c | 202a | 549a | |
| 6 | 131bc | 152a | 151bc | 135a | 446a | |
| 40 | 2 | 127bc | 163a | 135bc | 306a | 433a |
| 4 | 111c | 133a | 158bc | 182a | 415a | |
| 6 | 113bc | 118a | 115c | 133a | 234b | |
| P value | ||||||
| Temperature (A) | 0.0011 | 0.2624 | 0.0357 | 0.3659 | 0.0012 | |
| Period (B) | 0.6450 | 0.4359 | 0.5793 | 0.0319 | 0.1277 | |
| A × B | 0.3074 | 0.2607 | 0.1778 | 0.0057 | 0.2145 | |
운송기간 중 환경조건은 운송단계에서 아직 발달되지 않은 (육안으로 볼 수 없는) 화방의 과실 발육이나 수량 에는 영향을 주지 않으나, 발달되어 (육안으로 볼 수 있 는) 개화 전후 단계에 있는 화방의 과실 발육이나 수량 에는 영향을 미치는 것으로 보고되고 있다. Kubota와 Kroggel(2006)는 1화방(9-15mm 크기)이 육안으로 확인 가능한 단계의 토마토 묘를 4일간 운송하였을 때, 운송 중 암상태의 고온 조건에 놓인 토마토묘는 정식 후 1화 방의 꽃이 퇴화되거나 기부쪽1번과의 발육이 지연되어 1화방의 수량이 감소하였다고 보고하였다. 또한 6°C 또 는 13°C의 저온과 약광조사 조건에서는 이러한 1화방 꽃의 퇴화나 수량감소가 경감되었다고 보고하고 있다. 본 실험에서도 1화방 1번과의 무게가 40°C의 고온 처리 에서 감소하는 경향을 보여, 운송 중 온도 조건이 정식 후 토마토묘의 생육 및 수량에 영향을 미치며, 특히 운 송 중 고온 조건에 오랜 시간 있을 경우 수량의 감소를 초래하는 것을 확인할 수 있었다.
이러한 1화방 1번과의 무게감소는 개화 전후 고온에 의한 세포분열이 영향을 받은 데 기인한 것으로 추정된 다. Fanwoua 등(2012)에 의하면, 고온은 과실 생장 기간 을 단축시켜 최종 과실의 크기를 감소시키며, 이러한 감 소는 특히 세포분열이 일어나는 과실생장 초기 단계에 그 영향이 더욱 크다. 운송 온도처리시 토마토 묘는 1화 방의 1번화가 피기 직전 또는 핀 상태로, 이 시기의 고 온이 1화방 1번과의 과실생장에 영향을 미쳐 과실 크기 가 감소한 것으로 판단된다.
따라서 토마토묘의 운송시에는 온도조건이 지나친 고 온 또는 저온이 되지 않도록, 단순 상온유통이 아닌 온 도조절 장치 구비를 통한 온도조절이 요구된다. Kubota 와 Kroggel(2006)의 보고와 같이 4일의 기간이 걸리는 장거리 운송시에는 19°C 상온 조건으로 관리할 경우 정 식 후 토마토 꽃의 퇴화, 착과량 및 수량 감소 등이 초 래되므로 저온(6-13°C) 및 약광 조사(12μmol·m-2·s-1 PPF)가 요구되나, 6시간 이내의 단거리 운송시에는 10- 25°C 범위로 온도를 관리해줌으로써, 정식 후 장애 발생 없이 안정적인 착과 및 수량확보가 가능할 것으로 판단 된다.
