

서 론
재료 및 방법
1. 식물 재료 및 실험 방법
2. 생육조사
3. 성분 분석
4. 통계분석
결과 및 고찰
1. 야파처리에 따른 생육 분석
2. 기관별 RA, tilianin, acacetin 함량
서 론
배초향(Agastache rugosa Kuntze)은 향신료로 음식에 독특한 향과 풍미를 더 하기 위해 사용되는 꿀풀과의 허브이다(Do 등, 2020). 배초향은 에센셜오일, rosmarinic acid(RA), tilianin, acacetin과 같은 폴리페놀, 트리테르펜 등의 이차대사산물을 함유하고 있다(Han 등, 1987; Kim 등, 2018). 잎은 생으로 사용하거나 말려서 향신료로 사용할 수 있으며, 잎과 꽃에서 기능성 물질을 추출하여 식품, 의약품, 화장품 등으로 사용하고 있다(Kim 등, 2018). 복통, 설사, 구토 및 위장질환 등에 탁월한 효과가 있으며, 해열, 항균, 항암 작용에 효과가 있다고 알려져 있다(Shin, 2004; Wang 등, 2009; Zielinska과 Matkowski, 2014; Lam 등, 2019). 최근 건강 기능성 식품 및 화장품 산업이 커짐에 따라 배초향의 소요가 꾸준히 증가하고 있지만, 노지에서 재배 공급되는 배초향은 계절과 작기에 따라 생산량이 제한되고 제품 산업의 원료로 사용하는데 어려움이 있다. 특히 배초향은 노지 관행재배를 할 경우 연중 2회 수확이 가능하다고 보고되었으나 수요 증가에 따른 고품질 배초향의 공급량을 늘릴 필요가 있다(Rao와 Ravishankar, 2002).
식물공장은 노지재배의 단점을 극복할 수 있는 재배시스템으로, 폐쇄된 공간에서 적정 환경을 유지하여 작물의 생산성 및 균일성을 높일 수 있는 장점이 있다. 배초향을 식물공장에서 재배할 때 환경 제어를 통하여 안정적인 생산이 가능하며, 환경 제어를 통한 이차대사산물 함량을 증가시킬 수 있으므로 의약 원료로서 생산성과 품질을 향상 시킬 수 있는 배초향 재배기술이 필요하다(Kozai, 2018; Kim 등, 2020). 광과 온도, 습도, 배양액 등 식물의 생장 및 이차대사산물에 영향을 미치는 다양한 환경요인이 있다(Kim과 Lieth, 2003; An 등, 2017; Jung 등, 2017; Oh 등, 2017). 식물의 생육에 영향을 주는 광 관련 인자로는 광도, 광질, 광주기가 있는데, 그중 광주기는 식물의 개화와 생장, 형태형성 및 이차대사산물 함량에 영향을 미치는 중요한 요소이기 때문에 최적화가 매우 중요하다(Sugumaran 등, 2013; Cha 등 2014; Sul 등, 2022). 식물에 주어진 일장이 한계일장보다 길어지면 장일식물(long day plant, LDP)의 개화가 진행되고, 반대로 짧아지면 단일식물(short day plant, SDP)의 개화가 촉진된다(Thomas와 Vince-Prue, 1997; Runkle과 Heins, 2006). 장일처리 방법에는 명기가 종료되는 시점에 저조도의 광을 사용하는 명기 연장(day extension), 암기 중간에 빛을 공급하여 연속된 암기를 끊는 야파(night interruption)처리가 있다(Kim 등, 2015a). 야파처리는 식물의 장일 조건을 충족시켜 화훼류의 심비디움, 백일홍, 제라니움, 국화 등 장일식물의 개화 촉진과 생장 등에 효과가 있었으며, 오이에서 야파처리시 광합성을 증가시켜 과실의 품질을 향상시켰다(Trouwborst 등, 2010; Park 등, 2013; Kwon 등, 2013; Kim 등, 2015b).
야파처리를 이용한 식물의 개화 및 생육 조절에 관한 연구들이 주를 이루었으나 야파처리에 따른 이차대사산물의 증감에 관한 연구 보고는 많지 않다. 야파처리가 식물의 개화 및 생육 조절에 관여를 한다는 것은 각 기관별 이차대사산물의 생합성에 영향을 줄 수 있다는 것을 의미한다. 따라서, 본 연구는 식물공장에서 같은 DLI 조건에서 야파처리가 배초향의 생육 및 항산화 물질 함량에 미치는 영향을 이해하기 위해서 수행되었다.
재료 및 방법
1. 식물 재료 및 실험 방법
배초향 종자(Danong Co., Ltd., Korea) 240립을 240구 암면 배지(Kiemplug standard tray, Grodan, Denmark)에 파종하였다. 파종 후 온도 22 ± 2℃, 상대습도 70 ± 5%의 조건에서 형광등을 이용하여 광주기 20시간, 광도(PPFD) 180 ± 5μmol·m-2·s-1 조건 아래에서 5주간 재배하였다. 파종 2주 후 배초향 묘목을 EC 1.2dS·m-1의 호글랜드(Hoagland) 배양액에 매일 1분간 침지시켰다. 호글랜드 A배양액은 FeEDTA(4.898 g·L-1), Ca(NO3)2·4H2O(191.880g·L-1), KNO3(61.310g·L-1)를 사용하여 조제하였고, B배양액은 H3BO3(0.603g·L-1), MnSO4·5H2O(0.435g·L-1), ZnSO4·7H2O(0.044g·L-1), CuSO4 ·5H2O(0.010g·L-1), Na2MoO4·2H2O(0.004g·L-1), KNO3 (61.310g·L-1), MgSO4·7H2O(100.610g·L-1), NH4H2PO4 (23.470g·L-1)로 조제하였다. 본엽 4매가 전개되는 5주차에 각 처리당 8주씩 백색 LED(LED bar 40cm cool white, 10,000K, LG Innotek, Korea)를 사용하여 semi-DFT(Gafatec Co., Ltd., Korea) 시스템에 정식하였다. 정식 후 온도와 습도는 육묘환경과 동일하게 하였으며, 호아글랜드 배양액 EC 2.0dS·m-1, pH 6.5로 조절하여 공급하였다. 배초향의 생장 및 항산화 물질의 변화에 대한 야파처리 효과를 알아보기 위해 묘 군락 높이에서 광도 200μmol·m-2·s-1 조건으로 24시간 주기로 18:1:2:3, 18:2:2:2, 18:3:2:1(명기:암기:명기:암기) 비율로 06:00부터 조사하였으며 대조구는 20:4(명기:암기)로 하였다. 호글랜드 배양액을 사용하여 정식기간 동안 EC 1.8 ± 0.2dS·m-1, pH 6.5 ± 0.3으로 유지하였다.
2. 생육조사
정식 4주 후 배초향을 수확하였으며 지상부는 초장, 엽장, 엽폭, 지하부의 근장은 디지털 버니어캘리퍼스(SD500-300PRO, Shin Con Co., Ltd., Korea)를 이용하여 측정하였고, 화아의 개수를 조사하였다. 처리구별 엽록소 함량을 비파괴적으로 비교하기 위하여 SPAD-502(Minolta Camera Co., Ltd., Japan)을 이용하여 SPAD 값을 측정하였다. 지상부 생체중, 지하부 생체중은 전자저울(MW-2N, CAS Co., Ltd., Korea)을 이용하여 측정하였다. 건물중을 측정하기 위해 분리된 지상부와 지하부를 각각 시료분석용 봉투에 넣어 70℃로 설정한 건조기(HB-501M, Hanbaek Scientific Technology Co., Ltd., Korea)에 1주일간 건조시킨 후 동일 전자저울로 배초향의 지상부와 지하부 건물중을 측정하였다.
3. 성분 분석
배초향의 rosmarinic acid(RA), tilianin, acacetin 함량 분석을 위해 각각 꽃, 잎, 줄기, 뿌리로 나누어 시료분석용 봉투에 넣고 ‒70℃ 초저온 냉동고에 1일간 보관하였다. 이후 동결건조기(TFD5503, Ilshin BioBase, Korea)를 이용하여 4일 동안 완전히 건조시켰고, 완전히 건조된 배초향은 유발과 유봉을 이용하여 분말화하였다. 배초향 분말 0.1g을 2mg의 80% MeOH에 넣고 1시간 동안 초음파처리(sonication)를 실시 한 후 원심분리(12,000rpm, 10min, 4℃)하여 상등액을 추출하였다. 상등액을 0.45μm PTFE syringe filter로 여과한 다음 HPLC용 vial에 담아 HPLC(1260 Infinity, Agilent Technologies, Santa Clara, CA, USA)을 사용하여 분석하였다.
4. 통계분석
본 실험은 완전임의 배치법으로 수행되었으며 생육 및 기능성 물질 분석은 각 처리구별 3반복으로 실시하였다. 통계분석은 SPSS(Version 22.0.0.1, SPSS Inc., IL, USA) 프로그램을 이용하여 처리평균 간의 유의성 검증을 위해 Tukey의 다중검정법을 사용하였다(p ≤ 0.05).
결과 및 고찰
1. 야파처리에 따른 생육 분석
4주간 야파처리에 따른 배초향 생육 분석 결과, 엽장은 18:2:2:2와 18:3:2:1 처리구에서 유의하게 짧았고, 평균값으로 가장 짧았던 18:3:2:1 처리구는 대조구에 비해 8.82% 감소하였다(Table 1). 엽폭은 18:1:2:3 처리구에서 가장 길었고, 18:3:2:1 처리구보다 9.52% 증가하였다. 초장, 엽수, 근장, SPAD, 지상부와 지하부 생체중, 건물중은 유의적 차이를 보이지 않았다. 모든 야파처리구에서 대조구에 비해 화아가 증가하는 경향을 보였고, 18:1:2:3, 18:2:2:2, 18:3:2:1 처리구의 화아 수가 대조구보다 각각 16.95, 15.25, 22.03% 증가하였다. 이러한 결과는 단일 처리구의 배초향은 영양생장에서 생식생장으로 전환이 빠르게 진행되었다는 결과와 유사하였음을 알 수 있었다(Hwang 등, 2022). 배초향의 근장과 근중도 처음 암기가 길어질수록 비례하여 증가하는 경향을 보였다. 같은 꿀풀과에 속하는 바질도 일적산광량(day light integral, DLI)이 증가할수록 광합성에 의한 영양생장이 증가되지만, 동일한 DLI 조건에서는 생체중과 건물중에 영향을 미치지 않았다(Dou 등, 2018; Miyazawa 등, 2008). 본 연구에서도 유사하게 DLI가 14.4 mol·m-2·day-1로 동일하였기 때문에 모든 처리구에서 생체중과 건물중에 유의적 차이가 없었지만, 야파처리가 배초향의 잎의 형태형성과 개화를 촉진시키는 경향을 나타내었다.
Table 1.
Plant growth parameters of A. rugosa grown at four night interruption time (NIT) treatments (20:4, 18:1:2:3, 18:2:2:2, 18:3:2:1) at 28 days after transplanting.
| NIT |
Leaf length (cm) |
Leaf width (cm) |
Number of leaves | Flower buds |
Stem length (cm) |
Root length (cm) | SPAD |
SFW (g/plant) |
RFW (g/plant) |
SDW (g/plant) |
RDW (g/plant) |
| Control (20:4) | 7.4 az | 6.6 ab | 49.2 | 5.9 b | 40.6 | 57.4 | 59.1 | 15.9 | 6.2 | 2.1 | 0.5 |
| 18:1:2:3 | 7.2 ab | 6.9 a | 45.5 | 6.9 a | 38.3 | 50.1 | 58.4 | 16.0 | 5.8 | 2.4 | 0.5 |
| 18:2:2:2 | 6.5 b | 6.4 b | 56.0 | 6.8 a | 37.7 | 53.6 | 57.5 | 15.1 | 5.9 | 2.4 | 0.4 |
| 18:3:2:1 | 6.8 b | 6.3 b | 52.8 | 7.2 a | 39.2 | 54.7 | 60.2 | 16.0 | 6.1 | 2.5 | 0.5 |
| Significancey | * | * | NS | * | NS | NS | NS | NS | NS | NS | NS |
2. 기관별 RA, tilianin, acacetin 함량
배초향 꽃의 단위 건물중당 RA 함량은 18:1:2:3가 대조구 및 다른 처리구와 비교했을 때 유의적으로 낮은 값을 나타내었으며 18:1:2:3 처리구는 20:4, 18:2:2:2, 18:3:2:1 순서로 비교했을 때 27.9%, 26.4%, 20.9% 낮은 수치를 보였다(Table 2). 꽃 건물중당 tilianin과 acacetin 함량은 첫 번째 명기 이후 암기 시간이 길어질수록 유의적으로 높게 나타났고 18:3:2:1에서 가장 높은 함량을 보였다.
Table 2.
Rosmarinic acid (RA), tilianin, and acacetin concetration (mg·g-1 DW) in flowers, stems, leaves, and roots of A. rugosa subjected to night interruption time (NIT) treatments.
| NIT | RA (mg·g-1DW) | Tilianin (mg·g-1DW) | Acacetin (mg·g-1DW) | |||||||||||
| Flowers | Stems | Leaves | Roots | Flowers | Stems | Leaves | Roots | Flowers | Stems | Leaves | Roots | |||
| Contorl(20:4) | 3.31 az | 3.45 a | 2.44 c | 16.78 d | 3.54 b | 0.53 a | 2.75 b | 0.06 d | 1.37 c | 0.93 a | 1.59 c | 0.01 d | ||
| 18:1:2:3 | 2.58 b | 2.44 c | 1.59 d | 31.80 a | 3.28 b | 0.12 c | 2.73 b | 0.23 a | 1.92 b | 0.23 c | 1.70 b | 0.08 a | ||
| 18:2:2:2 | 3.26 a | 2.65 b | 3.47 b | 29.94 b | 4.17 a | 0.26 b | 2.70 b | 0.17 b | 1.97 b | 0.45 b | 1.20 d | 0.05 b | ||
| 18:3:2:1 | 3.13 a | 3.38 a | 5.94 a | 19.52 c | 4.29 a | 0.27 b | 3.13 a | 0.09 c | 2.14 a | 0.47 b | 1.76 a | 0.03 c | ||
| Significancey | * | * | * | * | * | * | * | * | * | * | * | * | ||
줄기 건물중당 RA는 20:4와 18:3:2:1에서 유의적으로 높았고, 가장 낮은 함량을 가진 18:1:2:3보다 20:4 처리가 41.39% 높은 함량을 보였다. Tilianin과 acacetin은 20:4 처리구에서 가장 높은 함량을 보였다. RA와 tilianin, acacetin 세 가지 모두 20:4 처리구에서 높은 함량을 나타냈고, 야파처리 후 첫 암기가 길어질수록 비례하여 함량이 증가하였다.
배초향 잎의 건물중당 RA 함량은 20:4 처리구를 제외하고 야파처리 후 첫 암기가 길어질수록 유의적으로 높아졌다. 가장 높은 수치를 나타낸 18:3:2:1 처리구의 잎 RA 함량이 대조구 및 가장 낮은 함량을 보인 18:1:2:3보다 143.44%, 273.58% 더 높았다. 잎의 건물중 당 tilianin 함량은 18:3:2:1 처리구가 대조구 및 18:1:2:3, 18:2:2:2 처리구와 비교했을 때 각각 13.8%, 14.7%, 15.9% 더 유의적으로 높았다. 건물중 당 acacetin 성분 함량은 18:3:2:1 처리구에서 유의적으로 높은 수치를 나타내었고 18:2:2:2 비율에서 가장 낮은 성분 함량을 보였다.
뿌리의 건물중당 RA 함량은 18:1:2:3 처리구에서 가장 높았으며 야파 이후 암기 시간이 증가함에 따라 증가하는 경향을 나타내었다. 가장 낮은 값을 보인 대조구에 비해 18:1:2:3 처리구의 RA 함량이 89.3%가 높았으며, 야파처리가 뿌리의 RA 함량 증가에 긍정적인 효과를 나타내는 것으로 판단된다. 건물중 당 tilianin의 함량은 RA와 유사하게 18:1:2:3가 유의적으로 높은 함량을 나타내었고, 첫 명기 이후 암기가 길어질수록 함량이 줄어들었다. 건물중 당 acacetin도 18:1:2:3 처리구에서 가장 높은 함량을 보였으며 tilianin과 동일한 경향을 나타내었다.
지상부의 이차대사산물 성분 분석 결과 건물중당 tilianin과 acacetin 함량은 줄기에서 가장 적었으며 광 환경에 많은 영향을 받는 꽃과 잎에서 상대적으로 높은 함량을 보였다. 폴리페놀류에 속하는 tilianin과 acacetin은 페닐알라닌(phenylalanine)에서 유래하며 페닐프로파노이드(phenylpropanoid)라고도 한다. 꽃과 잎 부위가 광에 노출되면 페놀 생합성 효소인 CHS(chalcone synthase)와 CHI(chalcone isomerase)가 tilianin, acacetin의 생산을 유도하고, 이후 줄기와 뿌리로부터 수송되어 꽃과 잎에서 생합성되기 때문에 tilianin과 acacetin 함량은 꽃과 잎에서 높게 나타났다(Dixon 등, 2002; Tuan 등, 2012; Yadav 등, 2020).
그리고 뿌리의 단위 중량당 RA 함량은 지상부 기관보다 높았다. RA의 생합성 과정에는 페닐프로파노이드 경로와 티로신 유도(tyrosine-derived) 경로를 모두 포함한다. 배초향이 광 스트레스를 받게 되면 뿌리의 티로신 유도에 속하는 유전자 전사체가 높은 발현량을 보이며 4-coumaroyl-CoA와 4-hydroxyphenyl-lactic acid를 생성하게 되고, 두 분자는 로즈마린산 합성효소(rosmarinic acid synthase)에 의해 RA가 생성되어 지하부에 축적된다(Petersen, 1997; Kwon 등, 2021).
배초향의 총 건물중당 RA함량은 18:1:2:3과 18:2:2:2에서 대조구보다 47.92, 51.46% 높았고, 건물중당 tilianin과 acactin은 18:3:2:1에서 유의적으로 높게 나타났다(Fig. 1). 건물중당 총 폴리페놀 함량에서는 18:2:2:2에서 가장 높은 함량을 보였다. 배초향의 항산화 활성은 꽃이 완전히 개화했을 때 가장 높은 이차대사산물의 함량을 보이고, 개화시기가 빠를수록 높은 항산화 함량을 가진다(Yeo 등, 2021). 개화를 촉진하기 위해 꿀풀과인 살비아에서 4시간 야파처리 시, 개화단축과 화아 개수가 증가되었다(Hong 등, 2014). 본 실험에서도 20시간을 연속으로 주는 것보다 18시간을 주었을 때 화아수가 유의적으로 증가하였고 개화 진행이 빠르게 진행되었기 때문에, 배초향의 폴리페놀도 대조구보다 높은 함량을 보였을 것으로 판단된다.
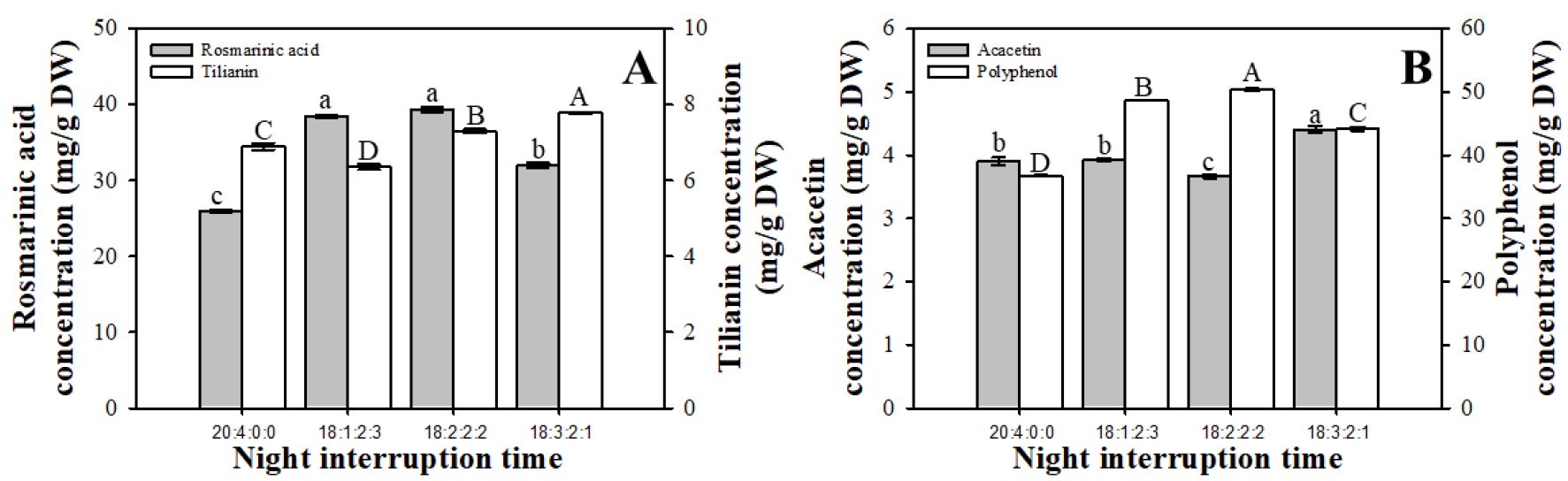
Fig. 1.
Rosmarinic acid, tilianin (A), acacetin, and pholyphenol (B) concentration of A. rugosa at different night interruption time treatments (20:4:0:0, 18:1:2:3, 18:2:2:2, 18:3:2:1). The data show the means and the vertical bars show the standard error (n=3). Different letters show significant differences among night interruption time treatments (Tukey’s test, p ≤ 0.05).
식물의 항산화 물질 중 하나인 페놀은 세포질의 경계면에서 합성되어 액포로 운반되어, 암기에는 이동을 멈추는 반면 연속광을 유지하면 페놀 함량이 증가한다(Engelsma, 1979). 광주기가 증가함에 따라 증가했던 페놀 함량은 암기가 길어지면서 분해되어 감소되며, 암기가 짧으면 페놀이 분해되기 전 축적되어 페놀함량이 증가한다(Engelsma, 1978). 본 실험에서 야파처리를 하였을 때 동일한 누적 광량으로 인해 생육 차이를 보이지 않았지만, 18:2:2:2의 야파처리가 총 폴리페놀 함량이 높게 나타났다. 따라서 인공광을 이용한 배초향 재배시스템에서 야파처리를 통하여 생육 저하 없이 배초향의 항산화 물질의 증대 효과를 기대할 수 있을 것으로 사료된다.
