

서 론
개량머루는 과피의 착색이 우수하고 내한성 및 환경 적응성이 뛰어나 생력 재배가 가능한 품종이다. 또한 당함량이 높고 신맛이 강하며, resveratrol과 같은 기능 성 물질이 풍부하여 술이나 음료와 같은 가공용으로 주 로 이용되고 있다(Kwon, 2012). 특히 지구 온난화로 인해 포도 변색기인 7~8월의 고온으로 착색이 불량해 지는 현상이 지난 수년간 발생되었기 때문에 외부 환경 의 영향없이 착색이 우수한 품종이 요구되고 있는 시점 에서 적합한 품종중 하나로 주목받고 있다. 하지만 화 진현상과 성숙기 탈립이 심하게 발생하고 과립이 1g 내 외로 다른 품종에 비해서 작아 상품성을 유지하거나 생 과로 이용하는데 문제가 되고 있다(Kim 등, 2004; Park 등, 2008).
생장조절제는 작물의 생장을 억제하거나 촉진하는 목 적으로 다양한 원예작물에서 이용되고 있다. 포도에서는 단위 결과 유도, 과립 비대, 착립수 증가 등을 목적으로 지베렐린이 이용되고 있고, 생리 활성을 가진 지베렐린 중에서 gibberellic acid(GA3)가 주로 이용되고 있다(Lee 등, 1996; Nickell, 1986; Park, 2010; Yamaguchi; 2008). 또한 합성 사이토키닌 thidiazuron(TDZ)과 chloropyridyl phenyl urea(CPPU)이 포도의 착립 증진 및 과립 비대에 효과 있는 것으로 보고되었다(Takahashi 등, 1978; Ogata 등, 1988; Tanakamaru, 1989).
따라서 본 실험은 개량머루의 과립비대 및 생산량 증 가를 위해 지베렐린과 사이토키닌을 처리하여 농도별 품 질 변화, 처리 적정농도 및 과피 조직의 해부학적 변화 를 구명하고자 수행되였다.
재료 및 방법
경기도 안성의 개인 농가에서 덕시설에 3.6m× 1.8m 로 재식된 8년생 개량머루 나무를 이용하여 실험을 수행 하였다. GA3 단독 처리시 적정 농도를 구명하기 위해 GA3 0, 50, 100, 200mg·L−1의 농도로 착립기에 각각 처리 하였다. 지베렐린과 사이토키닌의 혼용 처리 효과를 구명 하기 위해 GA3 단독 처리시 가장 성적이 우수했던 GA3 50mg·L−1과 100mg·L−1에 TDZ 0, 1.25, 2.5, 5mg·L−1을 각 각 혼합하여 무처리구와 비교하였다. 처리 방법은 만개 후 10일에 과방을 침지하였고, 각 처리구는 1주당 1반복으로 4반복 처리하였다. GA3(동부정밀화학)와 TDZ(N-phenyl-N- 1, 2, 3-thidiazol-5-yl urea, Bayer Cropscience)는 농업용으 로 시판되고 있는 제품을 구매하여 사용하였다.
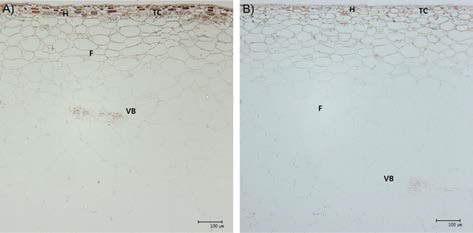
생장조절제 처리에 의한 과피 조직의 형태적 특성 변 화를 관찰하기 위하여 변색기와 수확기인 9월 15일과 10 월 5일에 각각 시료를 채취하였다. 시료는 과립의 적도부 위에서 과피 절편을 채취하여 2.5% glutaraldehyde, 1% osmium tetroxide에 세포를 고정하고, ethanol의 농도를 40, 60, 80, 90, 95, 100%로 단계적으로 높여 탈수 처리 하였으며, epon을 이용해서 포매하여 블록을 제작하였다. 이후 슬라이드 글라스에 ultramicrotome(PT-X, RMC, USA)으로 자른 박편을 치상한 후 PAS 염색법을 이용하 여 광학현미경(BX-51, Olympus, Japan)으로 관찰하였다 (Park, 1995).
과실 품질은 만개 후 108일에 일괄 수확하여 과방중, 과립중, 과립의 종경과 횡경을 측정하였고, 착즙된 과즙 으로 당도 및 적정 산도(0.1N NaOH, pH 8.1)를 조사하 였다. 과방중은 수확된 전량을 조사하였고 과립중은 주당 5과방을 선발하여 과립당 무게를 측정하였다. 통계처리는 PASW Statistics 18(IBM, US) 프로그램으로 Duncan의 다중검정을 실시하여 처리간 비교하였다.
결 과
GA3를 처리한 모든 처리구의 과방중(137.4-170.3g)이 무처리구(102.6g)보다 크게 생산되었고, 과방의 과립수는 75개인 무처리구에 비해 24.7-52.5% 증가하였다(Table 1). 과립중은 무처리구(1.26g)에 비해 17.5-32.5% 증가하였으 며, GA3 100 및 200mg·L−1처리구가 1.65, 1.67g으로 가 장 컸다. GA3 처리구의 과방이 착과수 증가와 과립 비대 로 인해 무처리구에 비해 과립이 밀착된 것을 육안으로 관찰할 수 있었다(Fig. 1). 당도 및 산도는 GA3 처리의 농도와 관계없이 무처리구와 차이가 나타나지 않았다.
Table 1.
Fruit characteristics and quality of Gaeryangmeoru grapes by GA3 treatment.
| Treatment (mg –1) | Cluster weight (g) | Berry/cluster | Berry weight (g) | Berry length (mm) | Berry diameter (mm) | Soluble solid content (°Brix) | Titratable acidity (%) |
|---|---|---|---|---|---|---|---|
| GA3 0 | 102.6 bz | 75.0 b | 1.26 c | 12.23 c | 12.11 c | 16.9 a | 1.67 a |
| GA3 50 | 145.0 ab | 101.0 ab | 1.48 b | 13.10 b | 12.89 b | 17.4 a | 1.57 a |
| GA3 100 | 170.3 a | 114.4 a | 1.65 a | 13.82 a | 13.90 a | 15.6 a | 1.66 a |
| GA3 200 | 137.4 ab | 93.5 ab | 1.67 a | 13.64 a | 13.80 a | 17.3 a | 1.77 a |

한편 GA3 처리는 고농도에서 과실에 약해를 유발하여 200mg·L−1 처리구에서 과실 외부에 검정 환형의 띠가 생 기는 과피 오염과가 발생되었고, 과방 끝부분에서 과경이 말려 올라가고 탈립되거나 소립과가 발생하였다(Fig. 2).

GA3 단독 처리 시험에서 과실 비대 효과와 약해 발생에 대한 안정성을 고려하여 GA3 50mg·L−1과 100mg·L−1에 TDZ를 혼용 처리하여 과실의 특성을 조사하였다(Table 2). GA3 50mg·L−1 처리구를 제외한 모든 처리구에서 무처 리구(97.9g)보다 과방중이 큰 과실이 생산되었다. 과립수 는 GA3 100mg·L−1과 TDZ 5.0mg·L−1 혼용 처리구(83.5립) 가 무처리구(79.3립)보다 약간 많았으나 다른 농도의 처리 구에서는 무처리구와 과립수가 유사하거나 적었다.
Table 2.
Fruit characteristics and quality of Gaeryangmeoru grapes by GA3 and TDZ treatment.
| Treatment (mg·L–1) | Cluster weight (g) | Berry/cluster | Berry weight (g) | Berry length (mm) | Berry diameter (mm) | Soluble solid content (°Brix) | Titratable acidity (%) |
|---|---|---|---|---|---|---|---|
| GA3 0 + TDZ 0 | 97.9 cz | 79.3ab | 1.08c | 11.29b | 11.51b | 19.0a | 1.27c |
| GA3 50+TDZ 0 | 94.1c | 62.8d | 1.38b | 12.67a | 12.62a | 18.0b | 1.36 bc |
| GA3 50+TDZ 1.25 | 121.8ab | 76.9ab | 1.41b | 12.46a | 12.59a | 18.3ab | 1.34 bc |
| GA3 50+TDZ 2.5 | 107.4bc | 70.2 bcd | 1.44ab | 12.30a | 12.52a | 17.0c | 1.42ab |
| GA3 50+TDZ 5 | 106.8bc | 65.3cd | 1.44ab | 12.36a | 12.62a | 16.7c | 1.47a |
| GA3 100+TDZ 0 | 110.7bc | 70.6 bcd | 1.44ab | 12.60a | 12.63a | 18.0b | 1.38ab |
| GA3 100+TDZ 1.25 | 115.9b | 72.6abcd | 1.47ab | 12.58a | 12.62a | 16.8c | 1.35 bc |
| GA3 100+TDZ 2.5 | 115.8b | 75.0abc | 1.48ab | 12.45a | 12.57a | 15.9d | 1.40ab |
| GA3 100+TDZ 5 | 135.1a | 83.5a | 1.53a | 12.65a | 12.79a | 15.3d | 1.48a |
과립중은 무처리구의 과실이 1.08g으로 가장 작았으며 GA3 단독 처리구와 TDZ 혼용 처리구에서 모두 과립 비 대 효과가 나타났다. GA3과 TDZ 처리 농도에 따른 과립중 은 GA3 50mg·L−1 처리구 중에서는 TDZ 2.5mg·L−1(1.44g) 와 5mg·L−1(1.44g) 혼용 처리에서 가장 컸으며 GA3 100mg·L−1 처리구 중에서는 TDZ 5mg·L−1 혼용 처리구 가 1.53g으로 가장 컸다. 동일한 TDZ 처리 농도에서는 GA3 100mg·L−1 처리구에서 과립중이 더 크게 생산되어 GA3 혹은 TDZ의 처리 농도가 높을수록 과립의 비대가 촉진되었다.
GA3과 TDZ 처리 농도별 과실 품질을 비교한 결과, 당도는 무처리구가 19.0°Brix로 가장 높았고 생장조절제 처리구의 과실은 낮은 수준이었다. GA3 50mg·L−1 처리 구 내에서는 TDZ 2.5mg·L−1와 5mg·L−1 혼용 처리구에서 과실의 당도가 가장 낮았다. GA3 100mg·L−1 처리구 중 에서는 GA3 단독 처리구가 가장 높고 TDZ 2.5mg·L−1와 5mg·L−1 혼용 처리구가 가장 낮았다. TDZ 농도가 동일 한 경우에는 GA3를 100mg·L−1으로 처리하는 것이 GA3 50mg·L−1 처리구보다 당도가 낮았다. 즉, GA3 혹은 TDZ 의 처리농도가 높을수록 과실의 당도는 낮았다. 적정산도 는 무처리구(1.27%)가 가장 낮았으며, GA3 50mg·L−1과 100mg·L−1 처리구내에서 혼용된 TDZ의 농도가 높은 처 리구에서 적정산도가 높았고 TDZ 농도가 동일한 경우 GA3의 농도에 의한 차이는 관찰되지 않았다.
GA3와 TDZ 처리에 의한 과실 조직의 변화를 광학현미 경을 이용하여 비교하였다(Table 3, Fig. 3). 개량머루의 과피층수는 6.3층이었고, GA3와 TDZ 혼용 처리구(5.7- 7.7)와 차이가 없었으며 표피조직에서 유관속조직까지의 세포층수에도 차이가 없었다. 반면 표피조직에서 유관속 까지의 길이는 무처리구(280.3um), GA3 50mg·L−1 단독 처리구(245.0um), GA3 50mg·L−1+ TDZ 1.25mg·L−1 혼용 처리구(319.7um)에서 가장 짧았으며, GA3 100mg·L−1 처 리구가 50mg·L−1 처리구보다 길었다. GA3 50mg·L−1 처 리구에서는 TDZ의 농도가 높은 처리구에서 과피 두께 가 두꺼웠으나 GA3 100mg·L−1 처리구내에서는 TDZ 농 도에 따른 두께의 차이의 경향이 뚜렷하지 않았다. 과실 에서 분리한 과피는 무처리구보다 생장조절제 처리구에 서 무거웠지만 농도에 따른 차이는 발견되지 않았다.
Table 3.
Changes of pericarp characteristics according to GA3 and TDZ concentrations.
| Treatment (mg L–1) | Cell layers of pericarp | Between epidermis and vascular bundle | Skin weight (g) | |||
|---|---|---|---|---|---|---|
| No. of cell layers | Thickness (µm) | |||||
| GA 0+TDZ0 | 6.3az | 16.3 | ab | 280.3 | c | 0.16c |
| GA3 50+TDZ0 | 7.7a | 15.3 | b | 245.0 | c | 0.18bc |
| GA3 50+TDZ 1.25 | 7.0a | 18.7 | a | 319.7 | c | 0.20ab |
| GA3 50+TDZ 2.5 | 7.3a | 16.3 | ab | 611.0 | ab | 0.22a |
| GA3 50+TDZ5 | 6.7a | 18.7 | a | 756.3 | ab | 0.20ab |
| GA3 100+TDZ0 | 5.7a | 16.3 | ab | 608.0 | ab | 0.19bc |
| GA3 100+TDZ1.25 | 7.7a | 16.8 | ab | 593.8 | b | 0.19bc |
| GA3 100+TDZ2.5 | 7.0a | 19.3 | a | 873.7 | a | 0.21ab |
| GA3 100+TDZ5 | 7.3a | 17.3 | ab | 648.0 | ab | 0.20ab |
고 찰
개량머루의 GA3 처리는 과립 비대에 있어 효과가 우 수하였다. GA3를 단독으로 처리하는 경우 100mg·L−1까 지는 농도가 높을수록 과립이 비대하였지만 그 이상의 농도에서는 차이가 없고 200mg·L−1처리구의 과실에는 과피오염과가 발생하고 과방이 말려 탈립 및 소과가 발 생하는 약해가 발생되는 경우가 있으므로 과립 비대를 촉진하기 위한 GA3 농도는 100mg·L−1 이하로 처리하는 것이 가장 효율적이었다. GA3와 TDZ를 혼용해서 처리 하는 경우 GA3와 TDZ 농도가 높을수록 과립 크기는 증가하였다. 거봉에서 GA3 단독 처리(Park, 2010)와 GA3 및 TDZ 혼용 처리(Lee 등, 2013)에서 과립 비대에 효과가 있다는 기존의 보고와 같이 개량머루에서도 효과 적이었다.
Youn 등(2000)과 Zhang 등(2007)은 동양배의 지베렐 린 처리는 당 조성에 영향을 미친다는 보고와 같이 GA3 처리로 인해 과실 품질이 영향을 받을 가능성이 있었지 만 개량머루에서는 GA3 처리에 의한 품질 하락은 발생 하지 않았다. 반면 지베렐린과 사이토키닌을 혼용 처리 한 결과 당도가 낮고 산도가 높은 과실이 생산되었고, 이는 거봉의 지베렐린과 사이토키닌 처리가 당도를 감소 시킨다는 Byun과 Kim(1995)의 보고와 일치하지만 지베 렐린과 사이토키닌 처리는 거봉의 당도를 증가시키거나 영향을 미치지 않는다는 보고와는 상반된 결과였다(Lee, 2013; Lee 등, 1996). Park(2010)이 지베렐린과 사이토 키닌 혼용 처리에 의해 수확시기가 지연된다고 보고한 바와 같이 지베렐린과 사이토키닌 혼용 처리에 의해 과 실의 성숙이 지연되어 성숙이 완료되지 못한 과실이 수 확되었기 때문이며, 지베렐린과 사이토키닌의 처리 농도 가 높을수록 수확이 지연되었다.
GA3의 단독 처리에서는 착립수가 증가하였으나, GA3 와 TDZ 혼용 처리에서는 착립수가 증가하지 않았다. 기 존의 연구 결과에서도 지베렐린과 사이토키닌의 혼용 처 리에 의해서 착립수가 증가되거나(Byun과 Kim, 1995; Lee 등, 1996) 증가되지 않은 상반된 결과가 보고되었다 (Nickell, 1986). Kwon(2012)에 의하면 개량머루는 착과 에서 초기 과립비대기까지 약 5일이 소요되어 그 기간 이 매우 짧아 생장조절제의 처리시기가 착립에 밀접하게 작용하기 때문에 착립 직후부터 탈립이 진행되기 전에 처리하는 것이 착립률을 높일 수 있을 것으로 생각되었 다. 반면 만개기의 생장조절제 처리는 무핵과를 생산하 고 과다한 착립은 압착에 의한 열과의 원인이 될 수 있 으므로 생육 단계에 맞게 처리 시기를 준수하는 것이 반드시 필요하였다.
생장조절제 처리가 과실의 세포에 미치는 영향에 대하 여 Matsui 등(1986)은 지베렐린 처리가 델라웨어 포도의 세포 비대를 촉진하고 shargal 등(2006)은 배의 사이토 키닌 처리가 세포 수를 증가시킨다고 보고되었으나 개량 머루의 GA3와 TDZ 처리는 과피의 세포층수에는 변화 없이 길이만 증가되어 세포분열에는 영향을 미치지 않고 세포비대만 유도되었다. 즉 생장조절제 처리에 의해서 과립 크기가 증가하는 것은 세포분열과는 관계없이 세포 비대에 의한 나타나는 현상임이 명확하게 구명되었다.
따라서 과립 비대 및 착립률 향상을 목적으로 생장조 절제를 처리하는 경우에는 착립 직후부터 2~3회에 나누 어 처리하는 것이 우수하고 균일한 처리 효과를 기대할 수 있다. 적정 처리 농도는 GA3 100mg·L−1 이하로 단독 처리하거나 GA3 50 + TDZ 1.25mg·L−1으로 혼용 처리하 는 것이 효율적일 것으로 판단되었다. 또한 개량머루는 GA3과 TDZ 혼용 처리에 의해 과실의 성숙이 지연되어 미숙과가 수확될 수 있으므로 충분히 성숙된 과실을 선 별하여 수확해야 할 것으로 생각되었다.
