

Introduction
Materials and Methods
1. Effects of DIF during seedling stage on tomato seedling quality
2. Effects of seedling-stage DIF on early yield after transplanting in greenhouse
Results and Discussion
1. Effects of DIF treatments during the nursery stage on tomato seedling quality
2. Effects of DIF treatments during the nursery stage on early growth and yield after transplanting
Conclusion
Introduction
Tomato (Solanum lycopersicum L.) is one of the most economically important greenhouse vegetable crops in East Asia, where the adoption of smart‑farm environmental control technologies and hydroponic culture systems has accelerated the expansion of long‑term cultivation systems (Kubota et al. 2008; Van Iersel and Gianino 2017). In such production systems, stable and uniform growth and development are essential for maximizing farmers’ income, because uniform early growth and stable establishment immediately after transplanting are critical determinants of overall yield potential. Poor early establishment often delays the development of the first inflorescence and reduces fruit set, ultimately resulting in substantial yield losses (Schwarz et al. 2010). Consequently, improving seedling quality during the nursery phase is a key management objective for securing productivity and economic returns in long‑term tomato cultivation. In greenhouse nurseries, thermal and light environments fluctuate substantially across seasons. During the early summer period (May to July), seedlings are frequently exposed to high temperatures, which exacerbate excessive stem elongation and lead to the production of weak transplants. High temperature and suboptimal light conditions reduce stem diameter, inhibit root system development, and limit leaf expansion, all of which contribute to diminished seedling quality (Bertram and Karlsen 1994; Myster and Moe 1995). With the increasing adoption of late-summer transplanting (e.g., August), followed by year-round harvest in large-scale Korean greenhouses, low-quality seedlings produced under high-temperature conditions have increasingly been associated with delayed post-transplant growth, slower inflorescence development, and reduced early yield. Thus, urgent need for environmental management strategies that maintain robust seedling quality under high-temperature stress becomes inevitable.
Manipulation of temperature regimes during seedling production has been widely evaluated as a non-chemical growth regulation strategy. Among these, the difference between day and night temperature (DIF) is recognized as an environmental factor for controlling stem elongation and improving transplant compactness (Bertram and Karlsen 1994; Langton and Cockshull 1997). In particular, a negative DIF (-DIF), in which night time temperature exceeds daytime temperature, strongly suppresses internode elongation and is considered a promising technique for mitigating heat-induced seedling elongation. The day–night temperature difference (DIF) influences the partitioning and storage of photosynthates by altering the balance between photosynthesis during the day and dark respiration at night, thereby regulating plant morphology and growth habit (Yamaura et al. 2023). In addition, short-term low- temperature pulses applied before sunrise have been shown to reduce stem elongation without increasing daily mean temperature, representing an energy-efficient approach for height control under warm conditions (Myster and Moe 1995).
Although numerous studies have evaluated the morphological responses of seedlings to DIF manipulation, research has predominantly focused on growth characteristics during the nursery stage (Ohtaka et al. 2020, Kim et al. 2023). Though studies like that of Li et al. (2015) studied yield characteristics based on DIF, far fewer studies have examined the carry-over effects of nursery temperature regimes on post-transplant vegetative growth, reproductive development, fruit set, and early yield performance (Blanchard and Runkle 2007; Ali et al. 2022).
Therefore, the objective of this study was to evaluate the morphological traits and chlorophyll content (as a physiological parameter) of tomato seedlings to differing DIF regimes during the nursery stage and to determine how variations in seedling quality influence early post-transplant growth, inflorescence development, fruit set stability, and early yield. By elucidating these carry-over effects, this research aims to provide science-based guidance for producing robust seedlings capable of withstanding high- temperature nursery conditions and to support the development of reliable transplant management strategies that enhance early reproductive stability and productivity in long-term greenhouse tomato cultivation.
Materials and Methods
1. Effects of DIF during seedling stage on tomato seedling quality
1.1 Planting material and growing conditions
A European-type tomato cultivar (Solanum lycopersicum L. ‘Redkhan’; Mifuko Seed Co., Ltd., Korea) was used in this study. Seeds were sown into 32-cell plug trays filled with a horticultural substrate (‘Fieldwang’; Nongwoobio Co., Ltd., Korea) composed of 69.9% cocopeat, 15% peatmoss, 10% perlite, and 5% zeolite (v/v). The nursery period was for 7 weeks from 10 April to 30 May 2025 in a fully controlled plant production system located within the Smart Farm Innovation Valley (Gyeongsangnam-do, Korea), using a growth chamber (EH-3600, Hi-point, Taiwan).
1.2 DIF treatments
DIF was defined as the temperature difference between the day and night periods. The photoperiod was maintained from 06:00 to 22:00, and the dark period from 22:00 to 06:00. Four DIF treatments were applied continuously throughout the 7-week seedling stage: -8°C DIF (18/26°C Day/night), -5°C DIF (20/25°C), 0°C DIF (22/22°C), and +5°C DIF (25/20°C). All growth chambers were operated under identical environmental conditions other than temperature, with relative humidity maintained at 70% and CO2 concentration held near ambient levels (approximately 420 μmol·mol-1). Irrigation was supplied via a bottom-watering system once every two days during the first 3 weeks of the nursery period. From week 4 to week 7, the irrigation frequency was increased to once daily to meet the increasing water demand of the seedlings and maintain stable moisture levels in the horticultural substrate.
1.3 Lighting conditions
Multi-wavelength LEDs (UV 385–396 nm, blue 425-450 nm, green 525 nm, red 625-660 nm, far-red 730 nm, and white) were used as the light source (Taiwan Hipoint corporation, Kaohsiung, Taiwan). Photosynthetic photon flux density (PPFD) was measured at centre and corners at the height of seedlings using a LI-180 spectrometer (LI-COR, Lincoln, USA). PPFD was maintained at 300 μmol·m-2·s-1 under a 16/8 h (light/dark) photoperiod. Environmental variables, including temperature, were continuously monitored and automatically adjusted to minimize fluctuations.
1.4 Growth Measurements
Seedling growth was evaluated weekly using nine plants per treatment (three per replication). Measurements included plant height, stem diameter, node number, leaf number, leaf length, leaf width, leaf area, SPAD value, and shoot and root fresh and dry weights.
Stem diameter was measured 1 cm above the substrate surface using a digital caliper (DC150P, Bluetec, China). Leaf length and width measurements were taken on the third fully expanded leaf below the apex. Leaf area was determined using a leaf area meter (LI-3100C, LI-COR, USA).
Fresh weights of shoots and roots were measured separately, then the samples were dried at 70°C for 72 h and reweighed to determine their dry weights. Chlorophyll content was estimated indirectly using a SPAD meter (SPAD‑502, Minolta, Japan), with three readings averaged per leaf.
1.5 Experimental design and statistical analysis
The experiment was conducted using a completely randomized design consisting of four DIF treatments with three replications. Each treatment included three 32-cell plug trays, for a total of 96 plants, and growth measurements were obtained from nine uniform plants selected across the three replications. All data were analysed using SAS 9.4 (SAS Institute Inc., Cary, NC, USA). Analysis of variance (ANOVA) was performed to test treatment effects, and Tukey’s honestly significant difference test was applied at P < 0.05 when significance was detected. Figures were generated using SigmaPlot 12.5 (Systat Software Inc., San Jose, CA, USA).
2. Effects of seedling-stage DIF on early yield after transplanting in greenhouse
2.1 Cultivation conditions
Seedlings produced under each DIF treatment were transplanted on 30 May 2025 into a plastic greenhouse located in Haman, Gyeongsangnam-do, Korea. Plants were grown for 13 weeks until 20 August 2025. Coir substrate (coconut coir) slabs (chip: dust = 5:5, v/v; Shinhan Atec, Korea) were used as the growing medium. Plants were spaced to achieve a density of 3.125 plants·m-2 (31 cm within-row spacing).
Greenhouse environmental control maintained average temperatures near 28 ± 2°C and 60-80% relative humidity. To maintain the internal temperature near 28 ± 2°C during the high-temperature period from late May to August, industrial air conditioners were operated. The cooling system was configured to activate automatically when the internal temperature exceeded 26°C, particularly during hours of high solar radiation from 09:00 to 18:00, to ensure a stable growing environment. A standard Yamazaki nutrient solution was supplied via drip irrigation with EC 2.8 dS·m-1 and pH 6.0. Irrigation was applied every 20 min from 07:30 to 16:30, delivering approximately 80 mL per event per plant.
2.2 Growth measurements
Vegetative growth parameters (plant height, stem diameter, node number, leaf length, leaf width, and stem diameter at the growth zone) were measured every 2 weeks. The growth- zone stem diameter was measured 10 cm below the inflorescence. Leaf area was estimated using a non-destructive regression model described by Carmassi et al. (2007):
Total leaf area per plant was calculated as the cumulative sum of estimated leaf areas based on sampled leaves. Reproductive development was evaluated by counting flower buds, open flowers, and fruit set for each inflorescence. Early yield was measured from the first two trusses; fruit number, fresh weight, and mean fruit weight were recorded. Yield per plant (g·plant-1) was converted to area-based yield (t·ha-1) using planting density.
2.3 Experimental design and statistical analysis
The experiment was arranged in a randomized complete block design with three replications, and seven plants per replication (21 plants per treatment). Growth and yield measurements were taken from three plants per replication (n = 9). All data were analysed using SAS 9.4 (SAS Institute Inc., Cary, NC, USA). Analysis of variance (ANOVA) was performed to test treatment effects, and Tukey’s honestly significant difference test was applied at P < 0.05 when significance was detected. Figures were generated using SigmaPlot 12.5 (Systat Software Inc., San Jose, CA, USA).
Results and Discussion
1. Effects of DIF treatments during the nursery stage on tomato seedling quality
1.1 Early versus late nursery stage: a reversal in growth pattern
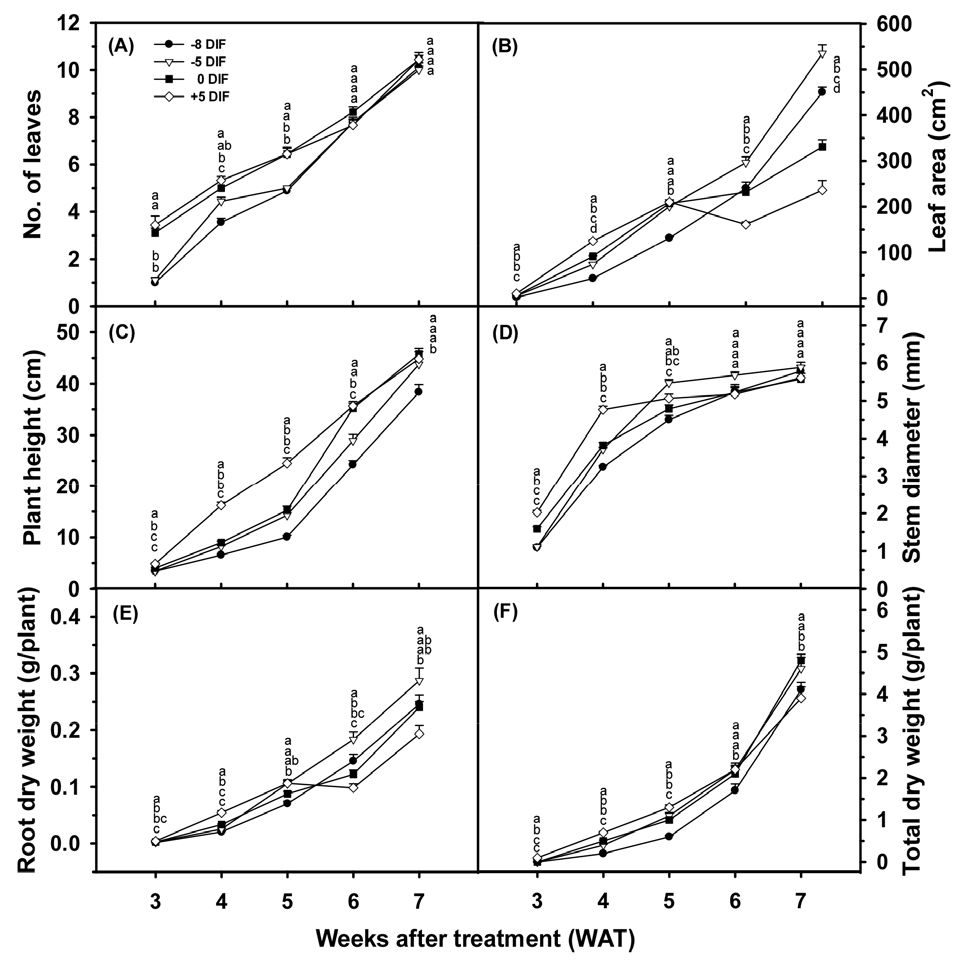
DIF treatments induced strong responses throughout the nursery period, but the treatment that performed best changed dramatically between early and late growth stages. During the first 3-4 weeks, seedlings grown under +5°C DIF showed the greatest leaf expansion, shoot elongation, and biomass accumulation (Tables 1, 2, 3). This reflects the well-known effect of higher daytime temperatures on photosynthetic activity and cell division (Grimstad 1994; Myster and Moe 1995; Moore et al. 2021). However, from week 5 onward, this pattern reversed. Seedlings under +5°C DIF developed leaf rolling, chlorosis, and premature senes-cence (Figs. 1 and 2), while those under -5 °C DIF, which had been slower initially, but gradually caught up and then surpassed all other treatments. By week 7, the -5°C DIF seedlings had achieved the largest leaf area (535.2 cm2, more than twice that of +5°C DIF seedlings), the thickest stems, the longest roots, and the highest total dry weight (4.6g) (Tables 2 and 3). This reversal shows that moderate negative DIF suppresses early elongation but preserves long-term physiological vigor, enabling sustained leaf and root development in the late nursery stage (Bertram and Karlsen, 1994; Ohtaka et al. 2020). Chlorophyll concentration (SPAD) initially favored +5°C DIF seedlings, but no treatment differences were detectable after week 5, confirming that DIF primarily affects quantitative growth traits (leaf expansion, biomass) rather than chlorophyll density.
Table 1
Effects of various DIF treatments on number of leaves, leaf area, leaf length, leaf width and SPAD values of tomato seedlings grown in a controlled growth chamber during the nursery periods
| DIF treatment | No. of leaves |
Leaf area (cm2) | Leaf |
SPAD values | ||
|
Length (cm) |
Width (cm) | |||||
| Growth stage: 3rd week | ||||||
| -8 | 18℃ / 26℃ | 1.0 bz | 1.8 c | 2.0 c | 1.2 c | 37.2 b |
| -5 | 20℃ / 25℃ | 1.1 b | 4.8 b | 2.4 c | 1.7 bc | 41.0 a |
| 0 | 22℃ / 22℃ | 3.1 a | 5.8 b | 3.1 b | 2.1 b | 38.1 b |
| +5 | 25℃ / 20℃ | 3.4 a | 9.9 a | 4.3 a | 3.4 a | 43.2 a |
| Growth stage: 4th week | ||||||
| -8 | 18℃ / 26℃ | 3.6 c | 43.1 d | 9.8 c | 7.5 c | 40.9 b |
| -5 | 20℃ / 25℃ | 4.4 b | 74.0 c | 12.4 b | 10.0 b | 38.6 b |
| 0 | 22℃ / 22℃ | 5.0 ab | 91.3 b | 12.3 b | 10.0 b | 42.4 ab |
| +5 | 25℃ / 20℃ | 5.3 a | 124.9 a | 16.0 a | 13.3 a | 45.9 a |
| Growth stage: 5th week | ||||||
| -8 | 18℃ / 26℃ | 4.9 b | 131.3 b | 15.2 b | 11.8 b | 43.9 a |
| -5 | 20℃ / 25℃ | 5.0 b | 200.3 a | 18.2 a | 16.0 a | 44.3 a |
| 0 | 22℃ / 22℃ | 6.4 a | 207.1 a | 18.7 a | 15.8 a | 48.6 a |
| +5 | 25℃ / 20℃ | 6.4 a | 210.4 a | 19.0 a | 15.4 a | 46.8 a |
| Growth stage: 6th week | ||||||
| -8 | 18℃ / 26℃ | 7.8 a | 239.8 b | 22.0 a | 18.3 a | 43.0 a |
| -5 | 20℃ / 25℃ | 7.8 a | 296.2 a | 22.4 a | 18.0 a | 42.5 a |
| 0 | 22℃ / 22℃ | 8.2 a | 231.4 b | 20.1 b | 18.9 a | 41.3 a |
| +5 | 25℃ / 20℃ | 7.7 a | 161.1 c | 18.0 c | 14.3 b | 41.9 a |
| Growth stage: 7th week | ||||||
| -8 | 18℃ / 26℃ | 10.1 a | 450.1 b | 24.5 a | 20.9 a | 45.6 a |
| -5 | 20℃ / 25℃ | 10.0 a | 535.2 a | 22.7 b | 17.7 b | 43.5 a |
| 0 | 22℃ / 22℃ | 10.4 a | 330.7 c | 22.0 b | 17.3 b | 45.1 a |
| +5 | 25℃ / 20℃ | 10.4 a | 235.9 d | 19.6 c | 16.9 b | 49.9 a |

1.2 Shoot elongation versus stem thickening and root growth
The temporal shift described above was accompanied by a fundamental change in plant structure. Under +5°C DIF, rapid internode elongation and node production occurred at the clear expense of stem thickening and root development (Table 2, Fig. 2). The result was weak, elongated shoots with relatively underdeveloped root systems which is a classic positive-DIF syndrome (Erwin and Heins, 1995; Kubota et al. 2008; Schwarz et al. 2010). In contrast, the -5°C DIF regime promoted only moderate early elongation, followed by steady stem thickening and robust root growth. By week 5, -5°C DIF seedlings already had the greatest stem diameter (5.5 mm) and longest root length (24.3 cm) (Table 2). Negative DIF is known to reduce gibberellin-mediated elongation and lower respiratory carbon loss under high daytime temperatures, thereby increasing carbohydrate availability for roots and supportive tissues (Song et al. 2023; Yamaura et al. 2023). By the end of the nursery period, the -5°C DIF seedlings had developed a compact, sturdy shoot–root architecture that closely resembles the description of high-quality transplants in earlier DIF studies (Myster and Moe 1995; Van Iersel and Gianino 2017). The +5°C DIF seedlings, by contrast, remained elongated and structurally compromised.
Table 2
Effects of various DIF treatments on number of nodes, stem diameter, plant height and root length of tomato seedlings grown in a controlled growth chamber during the nursery periods
| DIF treatment | No. of nodes |
Stem diameter (mm) |
Plant height (cm) |
Root length (cm) | |
| Growth stage: 3rd week | |||||
| -8 | 18℃ / 26℃ | 1.0 bz | 1.1 c | 3.4 c | 4.9 b |
| -5 | 20℃ / 25℃ | 1.0 b | 1.1 c | 3.4 c | 5.6 b |
| 0 | 22℃ / 22℃ | 2.0 a | 1.6 b | 4.0 b | 7.3 a |
| +5 | 25℃ / 20℃ | 2.0 a | 2.0 a | 4.8 a | 8.2 a |
| Growth stage: 4th week | |||||
| -8 | 18℃ / 26℃ | 3.6 c | 3.2 c | 6.6 c | 15.3 b |
| -5 | 20℃ / 25℃ | 4.4 b | 3.7 b | 8.2 b | 15.8 b |
| 0 | 22℃ / 22℃ | 5.0 ab | 3.8 b | 9.0 b | 17.6 ab |
| +5 | 25℃ / 20℃ | 5.3 a | 4.8 a | 16.2 a | 19.1 a |
| Growth stage: 5th week | |||||
| -8 | 18℃ / 26℃ | 4.9 b | 4.5 c | 10.1 c | 18.7 b |
| -5 | 20℃ / 25℃ | 5.0 b | 5.5 a | 14.3 b | 24.3 a |
| 0 | 22℃ / 22℃ | 5.2 b | 4.8 bc | 15.3 b | 21.6 ab |
| +5 | 25℃ / 20℃ | 6.0 a | 5.1 ab | 24.5 a | 22.2 a |
| Growth stage: 6th week | |||||
| -8 | 18℃ / 26℃ | 6.8 b | 5.2 a | 24.2 c | 21.8 a |
| -5 | 20℃ / 25℃ | 6.6 b | 5.7 a | 28.9 b | 24.7 a |
| 0 | 22℃ / 22℃ | 7.8 a | 5.2 a | 35.2 a | 22.6 a |
| +5 | 25℃ / 20℃ | 6.8 b | 5.2 a | 35.7 a | 22.6 a |
| Growth stage: 7th week | |||||
| -8 | 18℃ / 26℃ | 9.1 b | 5.8 a | 38.4 b | 23.1 a |
| -5 | 20℃ / 25℃ | 8.9 b | 5.9 a | 43.8 a | 24.7 a |
| 0 | 22℃ / 22℃ | 9.4 ab | 5.6 a | 45.7 a | 22.6 a |
| +5 | 25℃ / 20℃ | 10.0 a | 5.6 a | 44.8 a | 22.6 a |
1.3 Biomass accumulation
Fresh and dry weights also exhibited the same characteristic shift in treatment superiority between early and late nursery stages (Table 3, Fig. 2). During the early stage (3-5 weeks), seedlings grown under +5°C DIF had the greatest shoot and root fresh weights as well as the highest total dry weight, reflecting enhanced photosynthetic carbon assimilation and metabolic activity under higher daytime temperatures. However, by week 7, this pattern had reversed. Seedlings grown under -5°C DIF and 0°C DIF accumulated the greatest total fresh and dry weights. The -5 °C DIF treatment produced the highest total dry weight (4.6 g) and the greatest root dry weight (0.3 g) (Table 3), indicating superior biomass partitioning to the root system. In contrast, +5°C DIF seedlings exhibited a decline in total dry matter and a reduced proportion of biomass allocated to roots. This reduction supports the interpretation that positive DIF, while effective in accelerating early growth, may lead to premature senescence and diminished physiological vigor during extended nursery periods (Blanchard and Runkle 2007; Vaid et al. 2014). Further, An et al. (2021) reported that transplants with higher dry matter accumulation relative to plant height, and with well-developed root systems, exhibit superior early establishment and yield potential after transplanting. The seedlings produced under -5°C DIF in the present study consistently met these criteria, indicating that this DIF regime supports the development of structurally and physiologically robust transplants.
Table 3
Effects of various DIF treatments on fresh weight, and dry weight of tomato seedlings grown in a controlled growth chamber during the nursery periods
| DIF treatment |
Fresh weight (g/plant) |
Dry weight (g/plant) | |||||
| Shoot | Root | Total | Shoot | Root | Total | ||
| Growth stage: 3rd week | |||||||
| -8 | 18℃ / 26℃ | 0.2 cz | 0.0 b | 0.2 c | 0.0 c | 0.0 c | 0.0 c |
| -5 | 20℃ / 25℃ | 0.2 c | 0.0 b | 0.2 c | 0.0 c | 0.0 bc | 0.0 c |
| 0 | 22℃ / 22℃ | 0.3 b | 0.0 b | 0.3 b | 0.0 b | 0.0 b | 0.0 b |
| +5 | 25℃ / 20℃ | 0.6 a | 0.0 a | 0.6 a | 0.1 a | 0.0 a | 0.1 a |
| Growth stage: 4th week | |||||||
| -8 | 18℃ / 26℃ | 2.0 d | 0.2 b | 2.2 d | 0.2 d | 0.0 c | 0.2 c |
| -5 | 20℃ / 25℃ | 3.1 c | 0.2 b | 3.3 c | 0.3 c | 0.0 c | 0.4 b |
| 0 | 22℃ / 22℃ | 3.7 b | 0.2 b | 3.9 b | 0.4 b | 0.0 b | 0.5 b |
| +5 | 25℃ / 20℃ | 6.2 a | 0.4 a | 6.5 a | 0.6 a | 0.1 a | 0.7 a |
| Growth stage: 5th week | |||||||
| -8 | 18℃ / 26℃ | 5.6 c | 0.5 b | 6.1 c | 0.6 c | 0.1 b | 0.6 c |
| -5 | 20℃ / 25℃ | 9.7 b | 0.8 a | 10.5 b | 1.0 b | 0.1 a | 1.1 b |
| 0 | 22℃ / 22℃ | 10.0 b | 0.7 ab | 10.6 b | 0.9 b | 0.1 ab | 1.0 b |
| +5 | 25℃ / 20℃ | 12.9 a | 0.9 a | 13.8 a | 1.3 a | 0.1 a | 1.4 a |
| Growth stage: 6th week | |||||||
| -8 | 18℃ / 26℃ | 19.7 a | 1.1 ab | 20.7 a | 1.4 b | 0.1 b | 1.5 b |
| -5 | 20℃ / 25℃ | 22.2 a | 1.2 a | 23.4 a | 2.0 a | 0.2 a | 2.2 a |
| 0 | 22℃ / 22℃ | 21.3 a | 0.9 b | 23.2 a | 2.1 a | 0.1 bc | 2.2 a |
| +5 | 25℃ / 20℃ | 15.7 b | 1.0 b | 16.7 b | 2.2 a | 0.1 c | 2.3 a |
| Growth stage: 7th week | |||||||
| -8 | 18℃ / 26℃ | 35.9 ab | 1.2 b | 37.1 ab | 3.8 b | 0.2 ab | 4.1 b |
| -5 | 20℃ / 25℃ | 38.8 a | 1.5 a | 40.2 a | 4.4 a | 0.3 a | 4.6 a |
| 0 | 22℃ / 22℃ | 33.4 b | 1.2 b | 34.6 b | 4.5 a | 0.2 ab | 4.8 a |
| +5 | 25℃ / 20℃ | 21.6 c | 1.1 b | 22.7 c | 3.7 b | 0.2 b | 3.9 b |
2. Effects of DIF treatments during the nursery stage on early growth and yield after transplanting
2.1 Carry-over effects on vegetative growth after transplanting
Despite all plants being grown under identical conditions after transplanting, the DIF regimes imposed during the nursery stage produced clear carry-over effects on establishment, canopy development, and subsequent growth (Table 4, Fig. 3). During the first 3 weeks after transplanting, seedlings from the -5°C DIF treatment established the fastest canopy expansion and leaf development. At 1 week after transplanting, their leaf area (535.2 cm2) was more than twice that of +5°C DIF seedlings, and this advantage persisted through week 3 (Table 4). This rapid establishment directly reflected the superior root systems and balanced shoot structure that these seedlings had acquired during the nursery stage (Ohtaka et al. 2020). In contrast, +5°C DIF seedlings showed delayed leaf unfolding, thinner stems, and partial chlorosis symptoms consistent with excessive respiratory carbon consumption and poor root development during the nursery period (Langton and Cockshull 1997).
Table 4
Effects of DIF treatments during the seedling stage on leaf growth characteristics and SPAD values of tomato plants at different growth stages after transplanting in a greenhouse
| DIF treatment | No. of leaves |
Leaf area (cm2) | Leaf |
SPAD values | |||||
|
Length (cm) |
Width (cm) | ||||||||
| 1 week after transplanting | |||||||||
| -8 | 18℃ / 26℃ | 10.1 az | 450.1 b | 24.5 a | 20.9 a | 45.6 a | |||
| -5 | 20℃ / 25℃ | 10.0 a | 535.2 a | 22.7 b | 17.7 b | 43.5 a | |||
| 0 | 22℃ / 22℃ | 10.4 a | 330.7 b | 22.0 b | 17.3 b | 45.1 a | |||
| +5 | 25℃ / 20℃ | 10.4 a | 235.9 c | 19.6 c | 16.9 b | 49.9 a | |||
| 3 weeks after transplanting | |||||||||
| -8 | 18℃ / 26℃ | 12.9 ab | 3098.0 a | 29.4 a | 20.2 a | 51.6 ab | |||
| -5 | 20℃ / 25℃ | 13.8 a | 3350.5 a | 30.1 a | 20.0 a | 55.0 a | |||
| 0 | 22℃ / 22℃ | 12.0 bc | 2684.6 ab | 28.7 a | 19.5 a | 53.6 a | |||
| +5 | 25℃ / 20℃ | 10.8 c | 2554.4 b | 29.2 a | 19.9 a | 47.7 b | |||
| 5 weeks after transplanting | |||||||||
| -8 | 18℃ / 26℃ | 20.0 a | 4787.9ab | 29.7 b | 20.1 b | 53.3 a | |||
| -5 | 20℃ / 25℃ | 19.1 a | 4568.2b | 30.3 b | 18.6 b | 53.5 a | |||
| 0 | 22℃ / 22℃ | 19.7 a | 5933.6a | 39.5 a | 26.5 a | 54.3 a | |||
| +5 | 25℃ / 20℃ | 19.0 a | 5840.4a | 38.3 a | 26.2 a | 52.8 a | |||
| 7 weeks after transplanting | |||||||||
| -8 | 18℃ / 26℃ | 25.3 a | 5669.5 b | 27.2 b | 15.5 b | 52.0 a | |||
| -5 | 20℃ / 25℃ | 25.8 a | 6636.1 ab | 31.7 a | 24.4 a | 51.0 a | |||
| 0 | 22℃ / 22℃ | 25.4 a | 7411.7 a | 31.0 a | 20.5 a | 54.4 a | |||
| +5 | 25℃ / 20℃ | 24.6 a | 6818.4 ab | 28.0 ab | 15.7 b | 54.2 a | |||
| 9 weeks after transplanting | |||||||||
| -8 | 18℃ / 26℃ | 30.7 a | 6475.9 b | 25.7 a | 14.9 a | 55.0 a | |||
| -5 | 20℃ / 25℃ | 30.9 a | 7385.4 ab | 25.6 a | 14.2 a | 50.6 b | |||
| 0 | 22℃ / 22℃ | 31.1 a | 8111.0 a | 23.8 a | 13.4 a | 50.1 b | |||
| +5 | 25℃ / 20℃ | 30.0 a | 7662.5 ab | 26.4 a | 14.7 a | 52.1 ab | |||
| 13 weeks after transplanting | |||||||||
| -8 | 18℃ / 26℃ | 35.0 a | 6988.8 b | 23.8a | 13.6a | 62.6 a | |||
| -5 | 20℃ / 25℃ | 35.7 a | 7867.1 ab | 23.0a | 11.4b | 59.2 b | |||
| 0 | 22℃ / 22℃ | 35.3 a | 8600.0 a | 23.4a | 12.8ab | 60.4 ab | |||
| +5 | 25℃ / 20℃ | 33.0 b | 8000.6 ab | 23.6a | 12.6ab | 58.4 b | |||

During mid-growth (5-9 weeks after transplanting), plants from the 0 °C DIF and +5°C DIF treatments showed accelerated leaf area expansion, but this represented excessive vegetative dominance rather than balanced growth. Such over-vigorous vegetative expansion is known to divert assimilates away from developing fruit and reduce reproductive efficiency (Myster and Moe 1995; Si and Heins 1996). By the final evaluation at 13 weeks, plants from the 0 °C DIF treatment had the largest leaf area (8,600 cm2), but -5 °C DIF plants produced the greatest number of leaves (35.7 leaves), indicating sustained node production and stable vegetative development (Table 4). Chlorophyll concentration (SPAD) did not differ markedly among treatments throughout most of the growth period, although -8°C DIF plants showed the highest SPAD values by week 13, suggesting delayed leaf senescence under strong negative DIF.
2.2 Carry-over effects on stem growth and node differentiation
Persistent effects were also evident in stem elongation, thickening, and node development after transplanting (Table 5, Fig. 4). At 1 week after transplanting, seedlings from the +5 °C DIF treatment had the greatest number of nodes (10.0), reflecting accelerated meristematic activity during nursery production, but their stems were significantly thinner. In contrast, -5°C DIF seedlings maintained the thickest stem base (5.9 mm), demonstrating that the structural robustness developed during nursery production was retained after transplanting. This enhanced stem thickening under negative DIF has been attributed to reduced gibberellin- mediated elongation and more favorable carbon allocation to supporting tissues (Myster and Moe 1995).
Table 5
Effects of DIF treatments during the seedling stage on number of nodes, basal stem diameter, apical stem diameter, and plant height of tomato plants at different growth stages after transplanting in a greenhouse
| DIF treatment | No. of nodes |
Basal stem diameter (mm) |
Apical stem diameter (mm) |
Plant height (cm) | |
| 1 week after transplanting | |||||
| -8 | 18℃ / 26℃ | 9.1 bz | 5.8 a | 2.8 b | 38.4 b |
| -5 | 20℃ / 25℃ | 8.9 b | 5.9 a | 2.6 b | 43.8 a |
| 0 | 22℃ / 22℃ | 9.4 ab | 5.6 a | 2.8 ab | 45.7 a |
| +5 | 25℃ / 20℃ | 10.0 a | 5.6 a | 3.1 a | 44.8 a |
| 3 weeks after transplanting | |||||
| -8 | 18℃ / 26℃ | 12.9 a | 10.4 a | 3.9 b | 103.7 a |
| -5 | 20℃ / 25℃ | 13.7 a | 9.9 ab | 3.8 b | 101.1 a |
| 0 | 22℃ / 22℃ | 12.4 a | 9.1 bc | 3.7 b | 92.2 ab |
| +5 | 25℃ / 20℃ | 10.0 b | 8.1 c | 5.5 a | 80.1 b |
| 5 weeks after transplanting | |||||
| -8 | 18℃ / 26℃ | 19.8 a | 10.7 a | 6.3 b | 168.7 a |
| -5 | 20℃ / 25℃ | 18.9 ab | 10.9 a | 5.6 c | 161.3 a |
| 0 | 22℃ / 22℃ | 19.3 a | 10.9 a | 5.9 bc | 150.6 b |
| +5 | 25℃ / 20℃ | 18.2 b | 9.9 b | 7.0 a | 136.2 c |
| 7 weeks after transplanting | |||||
| -8 | 18℃ / 26℃ | 25.1 a | 10.8 a | 7.2 a | 221.8 a |
| -5 | 20℃ / 25℃ | 25.8 a | 11.0 a | 5.7 b | 209.1 b |
| 0 | 22℃ / 22℃ | 25.1 a | 11.1 a | 7.3 a | 198.8 c |
| +5 | 25℃ / 20℃ | 23.8 b | 10.1 b | 6.9 a | 186.3 d |
| 9 weeks after transplanting | |||||
| -8 | 18℃ / 26℃ | 30.4 a | 11.0 a | 6.0 a | 262.1 a |
| -5 | 20℃ / 25℃ | 31.1 a | 11.3 a | 5.4 a | 255.1 a |
| 0 | 22℃ / 22℃ | 30.8 a | 11.3 a | 5.9 a | 241.5 b |
| +5 | 25℃ / 20℃ | 29.2 b | 10.4 b | 6.0 a | 228.8 c |
| 13 weeks after transplanting | |||||
| -8 | 18℃ / 26℃ | 35.3 a | 11.2 a | 6.3 b | 269.5 a |
| -5 | 20℃ / 25℃ | 35.7 a | 11.3 a | 5.4 c | 270.1 a |
| 0 | 22℃ / 22℃ | 35.3 a | 11.4 a | 7.0 a | 263.7 a |
| +5 | 25℃ / 20℃ | 33.3 b | 10.6 b | 6.0 b | 259.8 a |
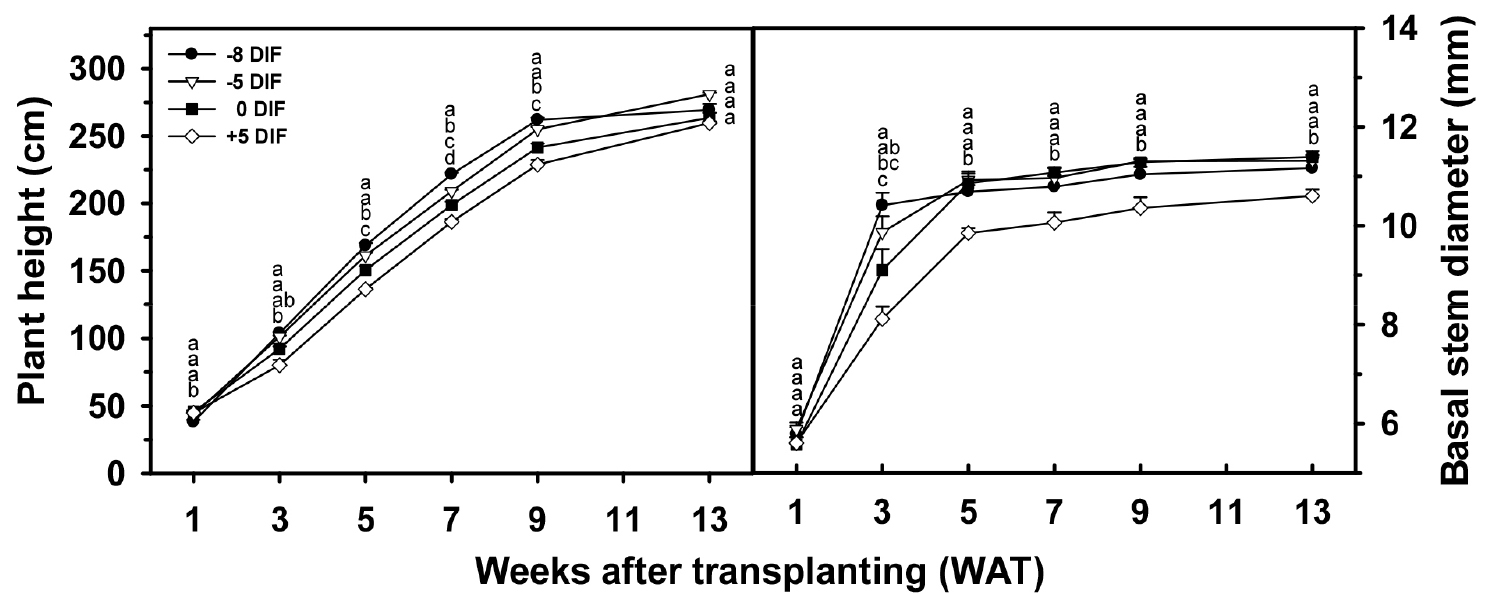
By week 3 after transplanting, both -5°C DIF and -8°C DIF seedlings exhibited greater plant height (approximately 101-104 cm) compared with the +5°C DIF treatment (80.1 cm) (Table 5). This pattern suggests compensatory growth, in which previously height-restricted seedlings increase elongation once environmental constraints are removed (Ohtaka et al., 2020). Notably, the -8°C DIF seedlings retained a base stem diameter substantially greater (approximately 28% larger) than that of +5 °C DIF plants, indicating that compensatory elongation did not compromise structural integrity. Throughout the remainder of the growth period, -5°C DIF plants maintained the most stable elongation rate and node formation, resulting in the greatest final plant height and node number at week 13 (Table 5). In contrast, 0°C DIF plants exhibited excessive apical thickening and vegetative dominance, while +5°C DIF plants consistently showed the lowest values for stem elongation, stem diameter, and node count. These outcomes align with previous reports that excessively high daytime temperatures during seedling production accelerate metabolic consumption of assimilates and negatively affect sustained growth after transplanting (Langton and Cockshull 1997).
2.3 Effects on flowering, fruit set, and earliness
Differences in seedling quality induced by DIF treatments during the nursery stage had pronounced carry-over effects on floral development, flowering, and fruit set after transplanting (Table 6). At 3 weeks after transplanting, seedlings from the -5°C DIF treatment produced the highest number of set fruits (2.1 fruits·plant-1), indicating superior early fruit-set stability. In contrast, −8 °C DIF seedlings produced the greatest numbers of floral primordia and open flowers, but their fruit set was limited (1.2 fruits·plant-1) (Table 6). This discrepancy suggests that excessive floral differentiation under strong negative DIF intensifies competition for assimilates, thereby reducing early fruit-set efficiency (Marcelis 1996; Kang et al. 2011). Seedlings from the +5 °C DIF treatment consistently showed the lowest numbers of flowers, open blooms, and set fruits, implying that positive DIF suppresses or delays the transition to reproductive development after transplanting.
During mid-growth (5-7 weeks after transplanting), plants in the -8°C DIF treatment displayed rapid progression of reproductive growth, producing the highest total numbers of flowers (28.0), open blooms (15.9), and fruits (9.1) by 7 weeks after transplanting. However, these values were not statistically different from those observed in the -5 and 0°C DIF treatments (7.8-8.0 set fruits) (Table 6), suggesting that enhanced floral initiation under strong negative DIF does not necessarily translate into improved fruit-set stability. This aligns with earlier findings that fruit retention depends more on physiological balance than on the sheer number of floral primordia (Marcelis 1996; Kang et al. 2011).
Table 6
Effects of DIF treatments during the seedling stage on lowers, number of blooms, and number of fruit sets of tomato plants at different growth stages after transplanting in a greenhouse
| DIF treatment | No. of flowers | No. of blooms | No. of fruit sets | |
| 3 weeks after transplanting | ||||
| -8 | 18℃ / 26℃ | 17.4 az | 4.2 a | 1.2 b |
| -5 | 20℃ / 25℃ | 13.8 b | 2.4 b | 2.1 a |
| 0 | 22℃ / 22℃ | 11.0 c | 1.7 b | 0.4 c |
| +5 | 25℃ / 20℃ | 8.1 d | 0.7 c | 0.2 c |
| 5 weeks after transplanting | ||||
| -8 | 18℃ / 26℃ | 24.1 a | 4.4 b | 8.7 a |
| -5 | 20℃ / 25℃ | 18.9 b | 8.7 a | 6.2 b |
| 0 | 22℃ / 22℃ | 22.4 a | 9.1 a | 6.9 b |
| +5 | 25℃ / 20℃ | 15.0 c | 8.2 a | 4.2 c |
| 7 weeks after transplanting | ||||
| -8 | 18℃ / 26℃ | 28.0 a | 15.9 a | 9.1 a |
| -5 | 20℃ / 25℃ | 25.1 b | 13.9 b | 7.8 a |
| 0 | 22℃ / 22℃ | 24.9 b | 11.6 c | 8.0 a |
| +5 | 25℃ / 20℃ | 24.1 b | 13.6 b | 5.3 b |
| 9 weeks after transplanting | ||||
| -8 | 18℃ / 26℃ | 28.2 a | 4.3 b | 9.1 b |
| -5 | 20℃ / 25℃ | 24.2 a | 3.2 b | 9.9 b |
| 0 | 22℃ / 22℃ | 26.2 a | 6.4 a | 13.7 a |
| +5 | 25℃ / 20℃ | 26.8 a | 3.6 b | 6.3 c |
Negative DIF treatments advanced harvest onset by at least 2 weeks compared with non-DIF or positive DIF conditions (Table 7). By 7 weeks after transplanting, approximately 55% of the plants in the -8 and -5°C DIF treatments had entered the harvestable stage, while no plants in the 0 or +5°C DIF treatments had produced ripe fruit (Fig. 6). At 9 weeks after transplanting, the -8°C DIF treatment showed the fastest transition to maturity, with 80% of plants having at least one harvestable fruit. However, full-plant harvestability was achieved simultaneously in -5 and 0°C DIF seedlings at 11 weeks after transplanting, indicating that strong negative DIF may accelerate early maturity at the cost of increased plant-to-plant variability. The -5°C DIF treatment ensured both advanced earliness and higher uniformity in harvest progression.
Table 7
Effects of DIF treatments during the seedling stage on cumulative fruit number, cumulative fruit weight, mean fruit weight, and estimated yield of tomato plants grown in a greenhouse
| DIF treatment |
Cumulative fruit numberz (no./plant) |
Cumulative fruit weight (g/plant) |
Mean fruit weight (g/fruit) |
Yield per planty (t/ha) | |
| -8 DIF | 18℃ / 26℃ | 6.9 ax | 657.6 ab | 95.6 a | 20.5 ab |
| -5 DIF | 20℃ / 25℃ | 7.6 a | 714.2 a | 98.9 a | 22.3 a |
| 0 DIF | 22℃ / 22℃ | 5.4 ab | 502.7 ab | 91.5 a | 15.7 ab |
| +5 DIF | 25℃ / 20℃ | 4.1 b | 406.3 b | 97.8 a | 12.7 b |
Cumulative yield per plant was highest in the -5°C DIF treatment (714.2 g·plant-1), approximately 75% greater than the yield obtained from +5°C DIF seedlings (406.3 g·plant-1) (Fig. 5). Estimated yield per hectare followed the same pattern (22.3 vs. 12.7 t·ha-1) (Table 7). Mean fruit weight did not differ among treatments (91.5-98.9 g), confirming that yield differences were attributable primarily to fruit-set stability and earlier harvest initiation rather than to differences in fruit growth rate or final fruit size. The high early yield in the -5 and -8°C DIF treatments can be attributed to robust seedling quality and stable floral differentiation achieved during the nursery stage. In particular, -5°C DIF facilitated balanced vegetative–reproductive growth, enabling more effective partitioning of assimilates to enlarging fruits (Heuvelink 1995, 1996; Marcelis 1996). Conversely, the low early yield in 0 and +5 °C DIF plants likely resulted from etiolation-like growth during the nursery stage, which increased vegetative assimilate consumption and reduced subsequent allocation to fruit sinks after transplanting (Talukder et al., 2025).
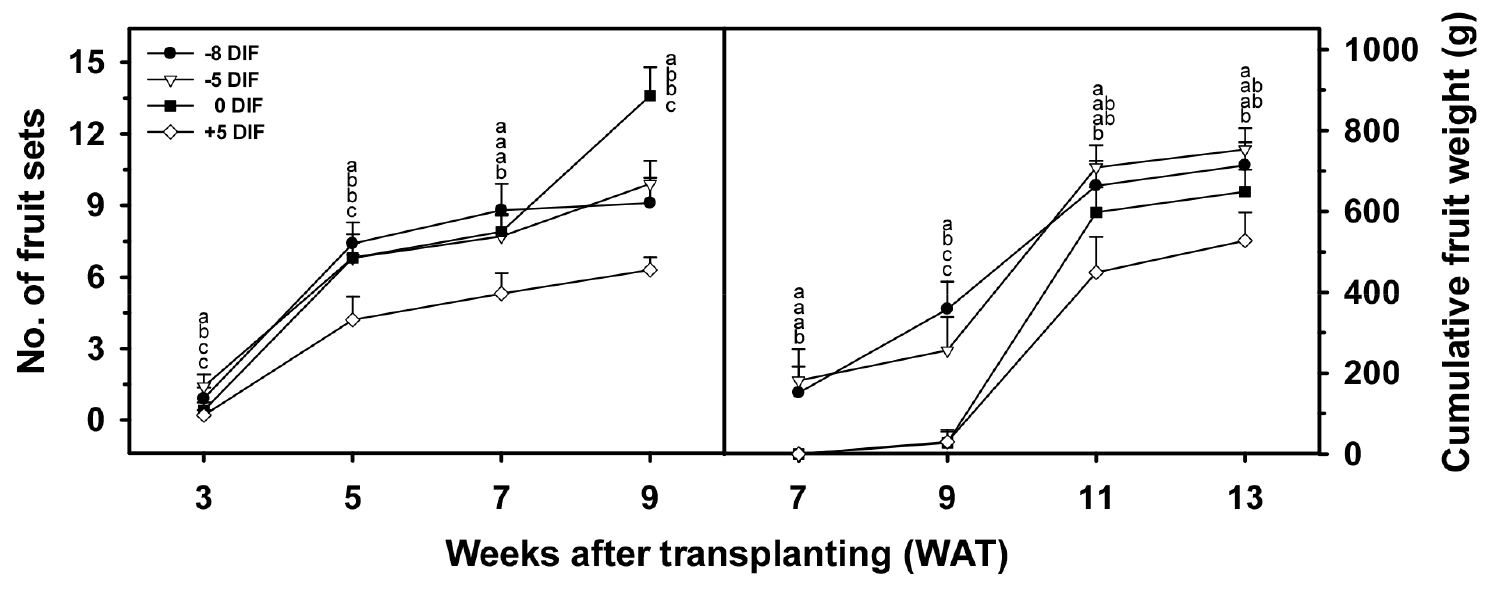
Fig. 5
Changes in number of fruit sets and cumulative fruit weight of tomato plants grown in a greenhouse as affected by DIF treatments during the seedling stage. Each parameter was assessed at 2-week intervals. Different letters are significantly different according to Tukey’s HSD test at P < 0.05 (n = 9)
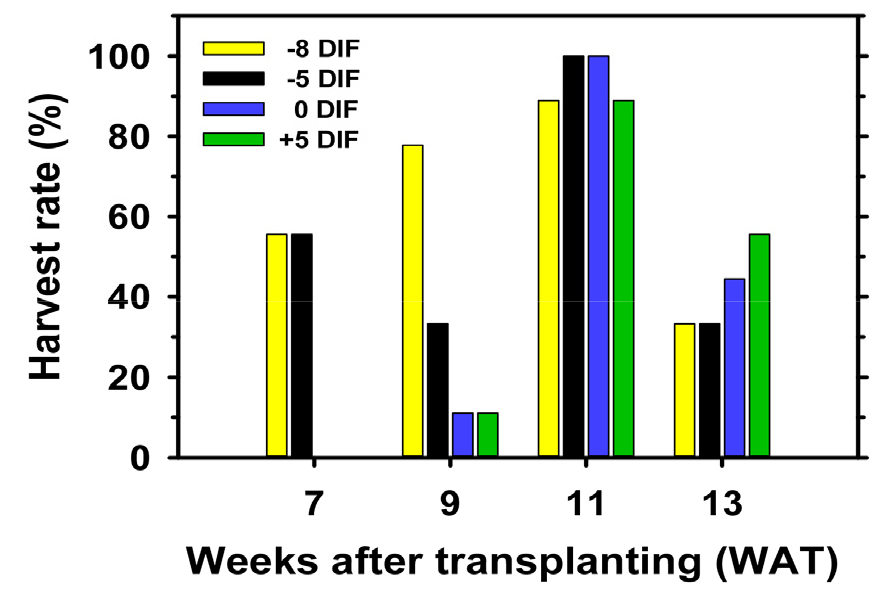
Fig. 6
Changes in harvest rate of tomato plants grown in a greenhouse as affected by DIF treatments during the seedling stage. Harvest rate was assessed at 2-week intervals and expressed as the percentage of plants producing harvestable fruits relative to the total number of plants per treatment (n = 9)
Taken together, the results demonstrate a consistent time-dependent pattern: +5°C DIF drives early rapid growth but leads to late-nursery senescence, poor root development, and post-transplant weakness, whereas -5°C DIF delays early expansion but produces compact, structurally balanced seedlings with superior late-nursery vigor, faster post- transplant establishment, higher fruit-set efficiency, and significantly greater early yield. These findings reinforce earlier reports that moderate negative DIF enhances transplant quality by regulating elongation growth without compromising later-stage vigor (Bertram and Karlsen 1994). Furthermore, Kim and Hwang (2019) demonstrated that precise environmental temperature control can produce high-quality, compact transplants without the need for chemical growth regulators, highlighting the potential of physical methods over chemical ones in modern transplant production systems. The present findings confirm that moderate negative DIF represents an effective method for managing transplant vigor and improving seedling robustness in high-temperature nursery environments.
Conclusion
This study demonstrated that differential day-night temperature regimes (DIF) applied during the tomato seedling stage exert significant and lasting effects on seedling quality, vegetative development after transplanting, reproductive performance, and early yield. Seedlings exhibited distinct growth responses depending on the magnitude and direction of DIF, confirming that thermal gradients act as strong morphological and physiological cues during early development. In contrast, -5°C DIF resulted in slower early shoot growth but produced seedlings with greater stem diameter, more balanced shoot-root development, and higher biomass accumulation by the end of the nursery period. These attributes are characteristic of high-quality, compact, and physiologically robust transplants. Carry-over effects of DIF treatments persisted after transplanting. Seedlings grown under -5°C DIF exhibited the most rapid recovery and canopy establishment, produced the largest leaf area and leaf number by 3 weeks after transplanting, and maintained superior stem strength and node differentiation throughout the growth period. Reproductive development was similarly influenced by nursery DIF conditions. The -5°C DIF regime produced the highest number of set fruits on the first truss and maintained stable fruit set throughout the cropping period, while -8°C DIF induced excessive floral initiation but resulted in low fruit-set efficiency. Overall, -5°C DIF represents the optimal thermal strategy for producing vigorous, compact tomato seedlings with superior post-transplant performance and early yield. Moderate negative DIF provides an effective, non-chemical approach for high-temperature nurseries.
