

서 론
도라지는 초롱꽃과(Campanulaceae)에 속하는 여러해살이풀로 한국, 중국, 일본에서 주로 재배되는 약용작물 중 하나이다(Lim, 1971). 도라지의 뿌리는 천연 기능성 물질인 사포닌 중에서도 Platycodin과 Platycodigenin의 함량이 높아 기관지 보호 효과가 우수하며 항비만, 항염증, 그리고 항암 효과 등이 있다(Ji 등, 2020; Zhang 등, 2020). 도라지의 뿌리는 한약재뿐만 아니라 식재료, 가공식품, 화장품 등 다양한 분야에서 이용되고 있으며(Jeong과 Yu, 2013; Park 등, 2011), 성분 추출과 같은 약리적 특성에 관한 연구가 활발히 이루어 지고 있다(Jung 등, 2022; Kim과 Jeong, 2015). 도라지는 건강기능식품의 관심이 높아짐에 따라 주요 증가가 기대되는 작물이다.
공정묘는 개별로 구분된 소량의 배지가 담긴 플러그 트레이의 셀에서 육묘하기 때문에 기존 직파방식의 육묘보다 빠르고 균일한 고품질 묘를 대량 생산할 수 있다. 공정 육묘는 시설 내에서 이뤄지는 육묘 방식으로 종자의 절약, 육묘 기간 단축을 통하여 비용을 절약할 수 있고, 자동화 및 기계화를 통해 노동력을 절감할 수 있다(Jeong 등, 2016). 현재 국내에서 공정 육묘에 대한 체계는 과채류 위주로 정립되어 있으며 약용작물의 공정 육묘 체계화는 부족한 실정이다(Jeong 등, 2016; Park 등, 2011). 기후변화로 인한 이상고온, 긴 장마로 인한 일조량 부족, 이상저온 등으로 인해 품질이 균일하지 않고, 대량 생산이 힘들기 때문에, 약용작물 공정육묘 체계를 도입할 필요가 있다(Bisbis 등, 2019).
공정 육묘 방식 중 하나인 폐쇄형 식물 생산 시스템(Closed- type plant production system, CPPS)은 계절과 장소에 대한 제약 없이 인위적으로 육묘에 최적인 온도, 습도, 광 등 환경을 조절할 수 있고, 이를 통해 육묘에 최적인 환경을 조성하여 출하 기간을 제어하고 육묘 기간을 단축할 수 있는 장점이 있다(Jeong 등, 2016). 또한, 다단식 육묘를 통해 토지 이용 효율을 증대시켜 균일한 고품질 묘를 대량 생산할 수 있으며, 정식 후 수확량 및 품질을 증대시킬 수 있다(van de Voren 등, 1986). 그러므로 고품질 공정묘의 생산을 위한 폐쇄형 식물 생산 시스템의 적정 환경을 구명하는 것이 중요하다(Johkan 등, 2010).
광 환경은 광도, 광주기, 광질 등 다양한 요소들로 구성되어 있다. 그중 광도와 광주기는 식물의 광형태형성, 성장 및 발달, 그리고 양분 흡수에 큰 영향을 주는 핵심 요소이다(Kang 등, 2013; Yan 등, 2019). 낮은 광도의 환경에서는 식물체의 순 광합성량이 감소하며 초장 및 절간장이 비정상적으로 늘어나는 도장이 발생한다(Fu 등, 2017; Yan 등, 2019). 반면, 광도가 증가하는 환경에서는 식물의 탄수화물 축적을 강화하고 순 광합성량을 증가시킬 수 있다(Dou 등, 2018; Lu 등, 2017). 하지만 광포화점 이상의 광도는 식물체의 기공을 폐쇄하여 광합성이 중단되고, 엽소 현상을 일으켜 작물에 피해를 일으킨다(Grime과 Jeffrey, 1965; Wimalasekera, 2019). 광주기는 식물의 생활환을 결정하는 중요한 요인으로, 식물의 영양생장과 생식생장을 조절한다(Jackson, 2009). 잎들깨와 같이 잎을 수확하는 단일 식물의 경우 전조를 통해 영양생장을 지속시켜 화아 분화를 억제한다(Chung과 Woo, 1988). 또한 광주기가 길어질수록 광보상점을 증가하고 광포화점에서의 순광합성량이 감소하는 경향이 있다(Carpenter 등, 1983). 광도와 광주기에 따른 식물의 반응은 다양하지만, 약용작물의 육묘를 위한 광도와 광주기에 관한 연구는 부족한 실정이다.
본 연구에서는 도라지 육묘를 위한 최적의 광 환경을 구명하고자, 다양한 광도와 광주기에 따른 도라지 묘의 지상부와 지하부 생육을 평가하였다.
재료 및 방법
1. 실험재료, 육묘 환경 및 광 환경 처리
도라지(Platycodon grandiflorum [Jacq.] A. DC. ‘Beak’, Asia Seed Co. Ltd., Seoul, Korea)는 4℃에서 2주 동안 저온 처리한 후, 50mg·L-1 NaOCl 용액으로 10분간 소독 후 10분간 흐르는 물로 헹궜다. 상업용 상토(Plant World, Nong Woo Bio, Suwon, Korea)로 충진된 50구 트레이(5 × 3 × 4.6cm; Bumnong Co. Ltd., Jeongeup, Korea)에 2024년 2월 22일에 소독된 종자를 파종하였다. 도라지는 온도 24.9 ± 0.9℃, 상대습도 53.7 ± 10.9%의 폐쇄형 식물 생산 시스템에서 48일간 육묘하였다. 양액은 온실다용도 액비[Ca(NO3)2·4H2O 472.5 mg·L-1, KNO3 252.5mg·L-1, KH2PO4 67.5mg·L-1, NH4NO3 40mg·L-1, MgSO4·7H2O 247.5mg·L-1, Fe-EDTA 11.1mg·L-1, H3BO3 1.3mg·L-1, MnSO4·H2O 1.0mg·L-1, CuSO4·5H2O 0.1mg·L-1, ZnSO4·7H2O 0.1mg·L-1, NaMoO4·2H2O 0.03mg·L-1]를 EC 0.7dS·m-1 및 pH 6.5로 맞추어 저면관수 하였다. 광질은 red:blue = 1:1 LED(Light emitting diode)를 이용하였다. 광도는 100, 150 그리고 200±10µmol·m-2·s-1 photosynthetic photon flux density(PPFD)로 설정하였고, 광주기는 10/14, 12/12, 14/10h(명기/암기)로 설정하였다. 광도는 트레이 높이에서 광도계(HD2102.2, Delta Ohm SrL, Caselle, Italy)를 이용하여 측정하였다.
2. 생육 조사
광도와 광주기에 따른 도라지 묘의 생육을 비교하기 위해 경경, SPAD, 엽면적, 그리고 지상부와 지하부의 생체중 및 건물중을 측정하였다. 경경은 버니어 캘리퍼스(CD-20CPX, Mitutoyo Co., Ltd., Kawasaki, Japan)를 사용하여 지제부 상단 2cm 지점을 측정하였다. SPAD 값은 엽록소 측정기(SPAD-502, Konica Minolta Inc., Tokyo, Japan)를 이용하여 완전히 전개된 잎 3개를 측정하여 평균값을 계산하였다. 엽면적은 엽면적 측정기(LI-3000, LICOR Inc., Lincoln, NE, USA)를 이용하여 측정하였다. 생체중 및 건물중은 전자저울(EW220-3NM, Kern&Sohn GmbH., Planegg, Germany)을 이용하여 측정하였다. 건물중은 70℃ 항온 건조기(Venticell-222, MMM Medcenter Einrichtungen GmbH., Planegg, Germany)에서 72시간 건조 시킨 후 측정하였다. 도라지 묘의 뿌리 발달 정도를 확인하기 위해, 지하부를 스캐너(Expression 12000XL professional scanner, Epson America Inc., Long Beach, CA, USA)로 취득한 이미지 데이터를 WinRhizo Pro 2020 뿌리 형태 분석 시스템(Regent Instruments Inc., Sainte-Foy, QC, Canada)을 이용하여 총 근장, 뿌리 표면적, 뿌리 부피, 근단 수를 측정하였다. 도라지 묘의 형태적 특성을 조사하기 위해 초장, 마디 수, 절간장 그리고 충실도(compactness)를 조사하였다. 초장은 지제부에서 식물체의 생장점까지의 높이를 측정하였다. 절간장과 충실도는 각각 Kim 등(2013)과 Jeong 등(2020)와 동일한 식을 이용하였다. 사용된 식은 다음과 같다.
3. 실험설계 및 통계분석
실험은 처리당 50개체씩 3반복의 완전임의배치법으로 배치하였으며 처리당 균일한 6개체의 묘를 채취하여 조사하였다. 통계분석은 SAS 프로그램(SAS 9.4, SAS Institute Inc., Cary, NC, USA)을 이용하여 분산분석(ANOVA)을 실시하였고, 평균 간 비교는 Duncan’s multiple range test를 이용하여 5% 유의수준에서 각 처리 간 유의성을 검증하였다. 그래프는 SigmaPlot 프로그램(SigmaPlot 14.5, Systat Software Inc., San Jose, CA, USA)을 이용하여 나타냈다.
결과 및 고찰
파종 후 48일 차 도라지 묘의 생육은 광도와 광주기가 증가할수록 도라지 묘의 경경, SPAD, 엽면적, 지상부와 지하부의 생체중과 건물중은 증가하는 경향을 나타냈다(Fig.1, Table 1). 특히, 100µmol·m-2·s-1에서 다른 광도와 비교하였을 때 광주기와 관계없이 도라지의 생육은 가장 저조하였고, 광주기가 짧아질수록 유의적으로 저조한 경향을 보였다. 150 µmol·m-2·s-1의 경우, 광주기가 길어질수록 경경, SPAD, 엽면적, 지상부와 지하부의 생체중과 건물중은 유의적으로 높아지는 경향을 보였다. 150µmol·m-2·s-1의 14/10h는 200µmol·m-2·s-1의 10/14h보다 일적산광량이 많았기 때문에, 경경, SPAD, 엽면적, 지상부의 생체중과 건물중 그리고 지하부의 건물중이 높았다. 도라지의 지상부 건물중은 200µmol·m-2·s-1, 12/12h에서 10/14h보다 유의적으로 높았고, 지하부의 건물중은 14/10h보다 유의적으로 감소시켰다. 도라지 묘의 엽면적은 모든 광도 처리에서 광주기가 10/14h일 때 유의적으로 좁았고, 12/12h와 14/10h 사이에 유의적인 차이는 없었다. 광합성은 작물의 광 에너지를 이용하여 탄수화물과 당 등을 합성하는 동화작용이다. 광합성을 통해 얻은 동화산물은 작물의 호흡 및 생육에 이용된다(Gardner 등, 2020). 광합성량은 광도가 증가할수록 증가하여 작물의 생육 및 생산성을 증가시킬 수 있다. 하지만 식물이 받는 광량이 광포화점 이상으로 증가하면 광합성량은 유의미하게 증가하지 않는다. 또한, 광주기가 길어질수록 광보상점이 높아지고, 순광합성량은 감소하는 경향이 있다(Carpenter 등, 1983). 그럼에도 불구하고 총광합성량이 증가하기 때문에, 작물의 생육이 향상될 수 있다. 그러므로 광포화점 아래에서 최적의 광도 및 광주기를 찾는 것이 중요하다. Fan 등(2013)은 낮은 광도에서 잎 면적당 기공 면적이 감소함에 따라 가스 교환량이 줄어들고, 이로 인해 광합성이 감소하여 작물의 생육과 생산성이 저해되며, 반대로 광도가 높을수록 기공의 수가 증가하여 작물의 생육이 우수하다고 보고하였다. 토마토와 상추의 경우 광도와 광주기가 증가할수록 순 광합성량이 증가하여 생육이 증가하는 경향을 보였다(Fan 등, 2013; Kang 등, 2013). 본 연구 또한 광도와 광주기가 증가할수록 작물의 생육이 우수해지는 경향을 보였고, 200µmol·m-2·s-1의 12/12와 14/10h에서 생육이 가장 우수하였다. 이는 총 광량의 증가에 따른 영향으로 판단된다.

Table 1.
The growth characteristics of Platycodon grandiflorum affected by different light intensities and photoperiods at 48 days after sowing (n = 6).
파종 후 48일 차 도라지 묘의 초장은 100과 150µmol·m-2·s-1에서 광주기가 길어질수록 길어지는 경향을 보였다(Fig. 2A). 200µmol·m-2·s-1의 12/12와 14/10h는 초장에 유의적인 영향을 미치지 않았다. 도라지 묘의 마디 수는 광도가 증가함에 따라 증가하는 경향을 보였다(Fig. 2B). 도라지 묘의 마디 수는 100µmol·m-2·s-1의 10/12h에서 가장 적었고 200µmol·m-2·s-1의 12/12h에서 가장 많았다. 또한 도라지 묘의 마디수는 150과 200µmol·m-2·s-1에서 10/14, 14/10, 12/12h 순으로 증가하는 경향을 보였다. 전체 초장 길이에서 마디 수를 나누어 계산된 절간장은 100과 150µmol·m-2·s-1에서 광주기가 길어질수록 길어지는 경향을 보였다(Fig. 2C). 하지만 200µmol·m-2·s-1의 12/12h에서 절간장은 다른 광주기보다 유의적으로 짧았다. 지상부의 건물중을 초장으로 나누어 계산한 충실도는 광도가 강해질수록 높아지는 경향을 보였고, 200µmol·m-2·s-1의 12/12와 14/10h에서 다른 처리보다 유의적으로 높게 나타났다(Fig. 2D). 광도와 광주기는 식물의 초장 및 절간장에도 큰 영향을 주는 요소이다. 옥신은 세포벽의 신장을 용이하게 하여 길이 생장을 촉진시킨다(Gardner 등, 2020). 하지만 약광 환경에서 옥신은 광분해 되지 않아 식물의 초장을 증가시킨다(Gardner 등, 2020). 반대로 광도가 높아질수록 초장 및 절간장은 짧아진다(Gardner 등, 2020). Chi(2003)는 토마토와 고추의 광주기를 단축시켰을 때 무기양분 흡수가 감소하고 황화현상이 나타났다고 보고하였다. 토마토와 오이 묘의 경우 광주기가 길어질수록 수분 흡수량이 증가하며, 이는 마디 수를 증가시키고 초장을 줄이는 경향을 보였다(Fan 등 2013; García-Caparrós 등 2020). 본 연구에서는 200µmol·m-2·s-1의 12/12h는 충실도가 높고 절간장이 짧은 도라지 묘 생산에 효과적인 광도와 광주기로 판단된다.
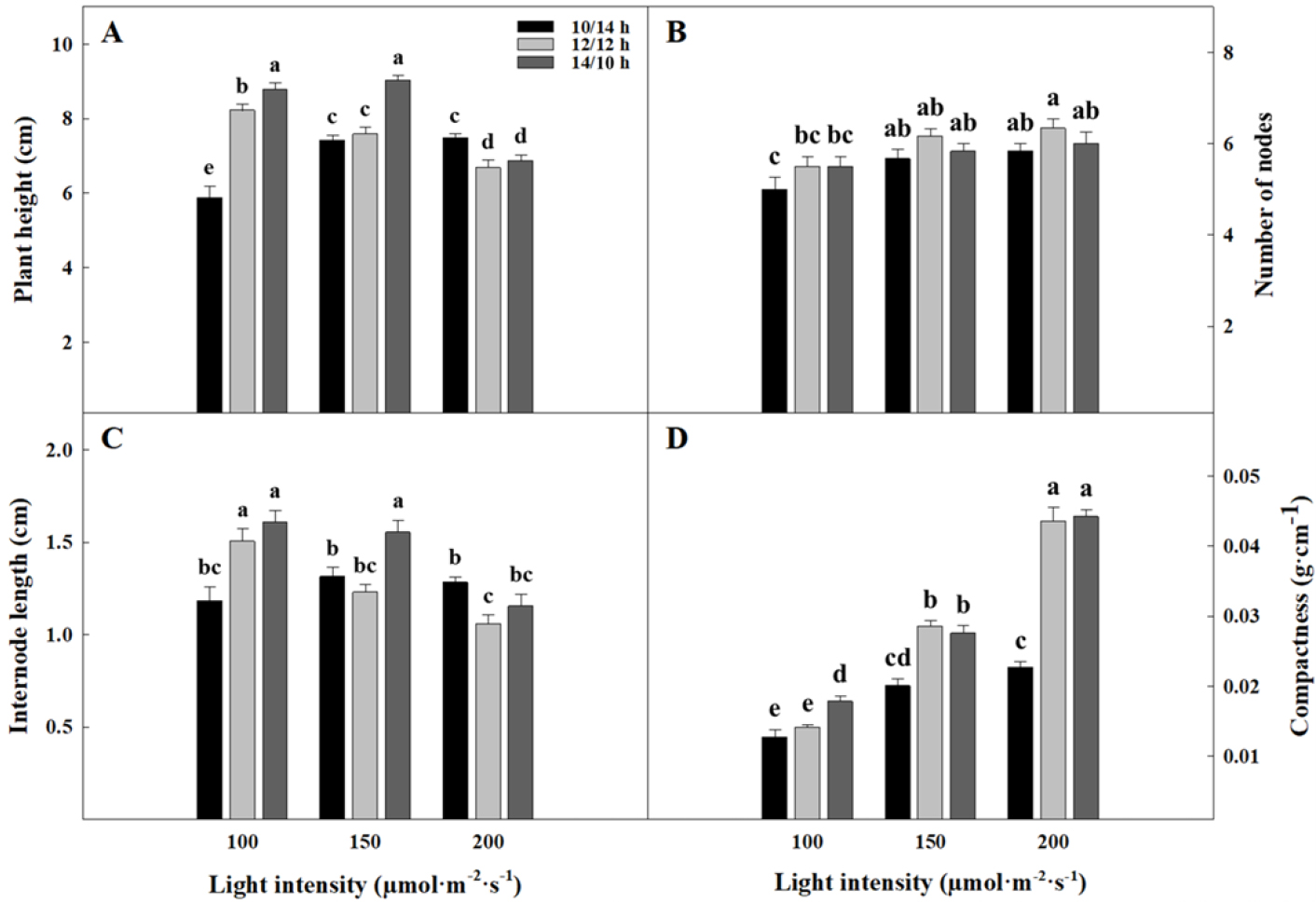
Fig. 2.
Plant height (A), number of nodes (B), internode length (C), and compactness (D) of seedlings of Platycodon grandiflorum as affected by different light intensities and photoperiods measured at 48 days after sowing. Vertical bars indicate standard errors of the means (n = 6). Different letters above bars indicate significant differences by Duncan’s multiple range test at p ≤ 0.05.
파종 후 48일 차 도라지 묘의 지하부 생육은 광도가 높아질수록 향상되는 경향을 보였다(Table 2). 특히 100µmol·m-2·s-1의 10/14h에서 뿌리 생육이 가장 저조하였다(Fig. 3). 150 µmol·m-2·s-1의 12/12h에서 총 근장과 뿌리 표면적은 다른 광주기보다 유의적으로 증가하였다. 특히 200µmol·m-2·s-1의 12/12h는 다른 처리보다 총 근장, 뿌리 표면적, 뿌리 부피 그리고 근단 수가 유의적으로 가장 높게 나타났다. 식물의 지하부 발달은 빛이 없는 지하에서 이루어진다. 하지만 뿌리의 형태형성은 지상부에서 광을 이용하여 생산된 식물 호르몬과 광합성 산물들이 영향을 미친다(Lee 등, 2017; van Gelderen 등, 2017). 지상부에서 생합성된 옥신과 자당은 지하부로 이동하여 지하부의 발달을 조절한다(Bhalerao 등, 2002; Kircher와 Schopfer, 2012; Reed 등, 1998). 광도와 광주기가 길어지면, 자당의 전달 및 호르몬의 작용이 증가하며, 뿌리의 성장, 양분 흡수와 바이오매스 축적을 촉진한다(Chen 등, 2016; Nagel 등, 2006). Guo 등(2019)은 광주기를 16/8h로 길게 하였을 때, 옥신 형성을 조절하는 ARF6 유전자의 발현과 뿌리 형성을 조절하는 WRKY2 유전자의 발현이 증가하는 것을 확인하였고 뿌리의 생육이 우수해지는 것을 확인하였다. Miotto 등(2021)은 광도와 광주기를 높일수록 주근의 길이가 길어지는 것을 확인하였다. 본 연구에서도, 광도가 가장 높은 200µmol·m-2·s-1에서 지하부 생육이 우수하였고, 광주기가 가장 짧은 10/14h에서 생육이 가장 저조하였다.
Table 2.
Root characteristics of Platycodon grandiflorum affected by different light intensities and photoperiods at 48 days after sowing (n = 6).
결론적으로 CPPS 내에서 도라지 묘를 생산할 때 광도와 광주기가 묘의 지상부 생육, 형태형성과 지하부 생육에 유의적인 영향을 주는 것을 확인하였다. 도라지 묘는 광도가 증가할수록 지상부 생육, 마디 수와 충실도가 증가하였고, 초장과 절간장은 단축되었다. 또한, 광주기가 길어질수록 생육이 향상되는 경향을 보였지만, 200µmol·m-2·s-1에서 12/12와 14/10h 사이에는 유의적인 차이가 나타나지 않았다. 지하부 생육의 경우 광도가 높아질수록 생육이 증가하였고, 지상부와 다르게 200µmol·m-2·s-1, 12/12h는 14/10h보다 생육이 우수하였다. 결과를 종합하여 봤을 때 CPPS의 광환경을 200µmol·m-2·s-1, 12/12h로 조성하면 충실도가 높고 생육이 우수한 묘를 얻을 수 있을 것이다.

