서 론
재료 및 방법
1. 생육조건 및 작물관리
2. 데이터 수집 및 생육측정
3. 흡광계수 측정 방법
4. 통계 및 데이터 분석
결과 및 고찰
결 론
서 론
오이(Cucumis sativus L.)는 온실 내에서 가장 많이 재배되는 작물 중 하나이며, 소득이 매우 높은 작물이다. 오이는 정식 후 약 40일 전후로 수확이 가능하며, 생장에 있어 온도와 광에 매우 민감한 작물이다(RDA, 2018). 일반적으로 온실에서 재배되는 작물은 광, 온도, CO2 등 환경요인에 영향을 많이 받으며, 특히 광은 동화산물생산의 필수 에너지원으로 생장뿐 아니라 수확량에도 영향을 준다. 온실내에서 작물이 직접 이용하는 광환경을 보다 정확하게 추적하고, 광 이용 특성을 이해하는 것은 매우 중요하다(Chen 등, 1999; Vos 등, 2010). 따라서 작물이 이용하는 광을 나타내는 수광형태에 대한 분석이 필요하다. 수광형태를 나타내는 지표 중 흡광계수는 군락내 높이에 따른 엽의 광 흡수율을 나타내며, 작물의 종류, 품종 그리고 재식밀도에 따라 차이를 보인다. 흡광계수의 정확한 측정을 통해 작물의 광이용형태에 대한 분석과 군락 내 개별 엽의 광합성 추정의 정확도를 높일 수 있다(Higashide, 2022; Nobel 등, 1993). 엽면적지수(leaf area index, LAI)는 흡광계수 측정의 주요한 변수로 수광량에 직접적인 영향을 주는 요인이다. Monsi와 Saeki(2005) 및 Higashide(2022)는 LAI가 증가할수록 높이에 따라 수광율이 변화한다고 하였으며, 이는 Lambert-Beer 법칙을 따른다고 보고하였다. 하지만 Tan 등(2020)과 Jones(1992)은 LAI뿐만 아니라 작물이 생장함에 따라 전개되는 잎의 각도, 신초의 생성 형태, 초장 등이 주변 작물의 광환경에 영향을 미칠 수 있다고 보고하였다. 온실내에서는 구조물의 위치와 태양고도 변화 때문에 생기는 그림자도 고려해야 하며, 또한 적엽과 같은 작물 관리는 군락내에 들어오는 광량을 변화시킨다고 보고하였다(Nederhoff, 1983; Nobel 등, 1993; Hirose, 2005). Peil 등(2002)은 추정 모델의 정확도를 높이기 위해서 흡광계수의 일별 변화량의 분석이 필요하다고 하였으며, 여름에 작물의 고온 피해를 방지하기 위해 사용하는 스크린이 확산광을 유도함에 따른 변화도 고려해야 한다고 하였다. 따라서 군락 내 광의 정확한 예측을 위해서는 작물의 생장과 온실 내에서 변화하는 광환경을 모두 반영하는 흡광계수에 대한 연구가 필요하다. 본 연구는 오이의 생육단계에 따라 연속적인 흡광계수를 측정하여, 작물의 생육 요소들과 흡광계수에 대한 관계를 분석하고, 생육단계를 반영한 오이 작물의 흡광계수 추정 모델 개발을 하고자 수행되었다.
재료 및 방법
1. 생육조건 및 작물관리
실험은 안동대학교 생명과학대학 연구온실(N36.5°, E128.8°, 4연동, 일중 피복, polyolefin film; 폭 25m; 길이 24m; 높이 9.6m; 측고 6m)에서 2022년 3월 24일부터 2022년 6월 30일까지 진행되었으며, 백다다기 오이 ‘Daemyung’(Wonnong Seeds Co., Ltd., Korea)을 사용하였으며, 육묘장으로부터 유묘를 구입하여 재배하였다. 정식 전 2주 동안 발근을 위해 암면큐브(KB101, UR Media, Korea)에 묘를 옮긴 후(10:00-15:00, 1시간 간격으로 500mL/day) 관수를 실시하였다. 큐브 하단부에서 발근을 확인 후 암면 슬라브(GT Master, Grodan, the Netherlands)에 정식을 하였다.
작물 관리는 덩굴손 제거, 측지 및 마디당 과실 1개를 유지하기 위해 주 2회 관리하였다. 또한 줄내림 재배 방식 및 유인을 위해 원줄기 재배를 하였으며, 초장이 220cm를 넘어가면 유인줄을 내려 지상으로부터 250cm를 넘지 않게 유인 관리하였다. 엽수는 노엽 제거와 작물 하부의 공기 순환을 위해 17매 수준을 유지 관리하였다. 슬라브당 작물은 3개체씩 정식하였으며, 재식밀도는 2.6주/m2였다.
여름철 시설 내 고온에 의한 작물의 해를 피하기 위해 포그 시스템과 차광스크린을 이용하여 환경관리 하였다. 실험기간 동안 시설내의 일평균 온도는 18-32℃, 상대습도는 80-97%이었다(Fig. 1). 관수는 적산일사량 기준 100J에 도달하였을 때 드리퍼를 이용하여 작물 1주당 80-120mL의 양액을 급액하였다. 양액의 조성은 야마자키 오이 표준 배양액을 사용하였다. EC는 정식 후 3주간 0.6에서 0.8dS·m-1까지 순차적으로 증가시켰으며, days after transplant(DAT) 30일 이후에는 1.2dS·m-1까지 높여주었다. 산도(pH)는 5.8-6.2 범위가 유지되도록 관리하였다.
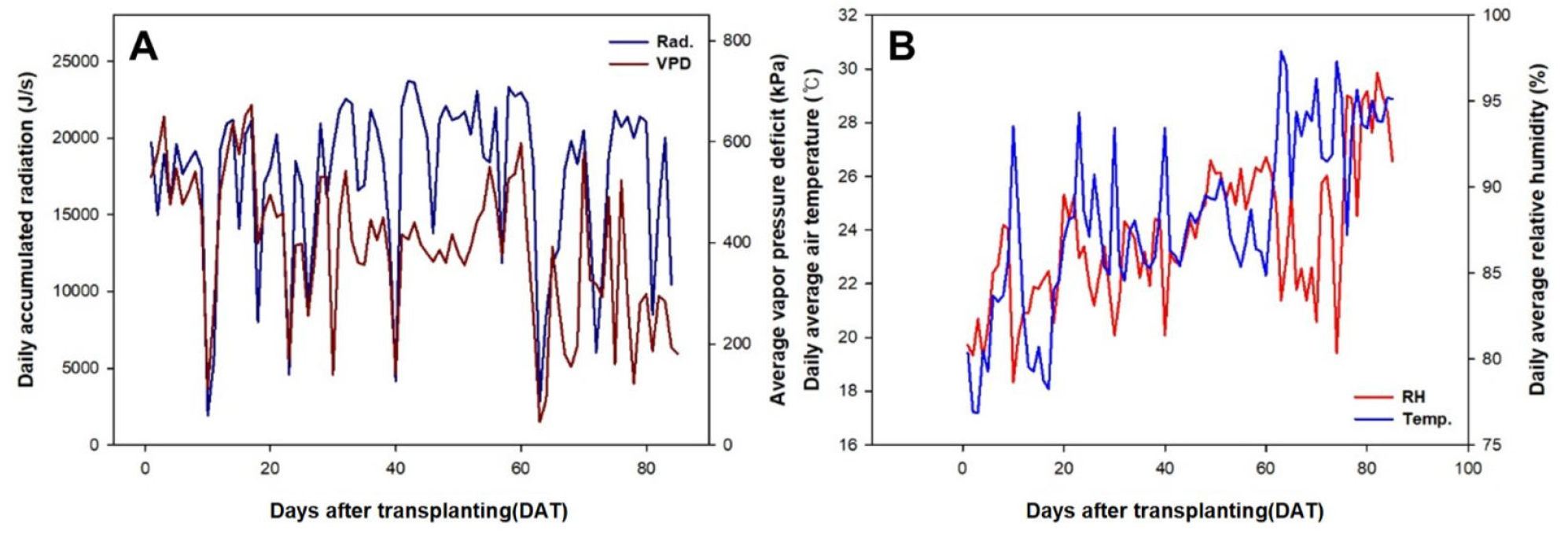
Fig. 1.
Changes in radiation, vapor pressure deficit (A), temperature, and relative humidity (B) during the experiment in the greenhouse.
2. 데이터 수집 및 생육측정
높이에 따른 광량은 작물의 초장을 5등분하여 각각 5곳의 위치에 일사량계(SP-110, Apogee, USA)를 설치하였다. 높이 별 광도는 데이터로거(CR1000, Cambell Scientific, USA)에 1초 간격으로 저장되었다. 군락 내 작물의 생장에 따른 상호간섭을 최소화하기 위해서 일중 누적 광량을 측정하여 비교하였다. 측정 개체의 선정은 수광 분포 특성 및 작물 간 상호간섭 파악을 위해 군락의 중심부에 있는 1개체를 임의로 선정하였으며, 측정 위치의 조절 후 3번 반복 측정하였다. 측정 장비의 위치는 3일 간격으로 일몰 후 광량이 더 이상 증가하지 않을 때 조정하였다(Fig. 2). 군락 내에서 임의적으로 6개체를 샘플링하여 일주일 간격으로 비파괴 측정을 하였다. 조사항목으로 초장, 엽수, 관부직경, 마디수를 측정하였다. 오이의 엽면적은 엽면적 측정기(LI-3000C Area Meter, LI-COR Inc., USA)를 이용하여 측정하였다. 흡광계수 측정 개체의 엽수와 초장이 동일한 1개체를 선정하여 7일 간격으로 순차적으로 파괴조사하였으며, 엽면적지수는 다음 식으로 계산되었다(Eq. 1).
* LAI: Leaf area index, LA: Leaf area D: Plant density

Fig. 2.
Experimental view (A) and diagram (B) for the measurement of light extinction coefficient.
3. 흡광계수 측정 방법
각각의 높이에서의 측정된 광량(I)은 최상단(I0)에서 들어오는 광량이 식물체에 의해 흡수된 후 투과된 광량을 의미하며, 각 위치에서 흡수된 광 비율을 통해 최종적으로 흡광계수를 구하였다. 흡광계수를 계산하기 위해서 Eq. 2과 Eq. 3을 이용하였으며, 각 높이에 따른 흡광비율을 모두 적분하여 일 누적 흡광비율을 구하였으며, 이를 DAT에 따른 LAIf로 나누어 일별 흡광계수를 계산하였다. LAIf는 DAT에 따른 변화를 피팅 후 일자에 맞는 값을 대입하였다.
* I: Light intensity in the canopy, I0: Light at the top of the canopy, k: Extinction coefficient, LAIf: Leaf area index
4. 통계 및 데이터 분석
엽면적지수(LAI)와 초장에 따른 흡광계수의 관계분석은 다양한 모델을 사용하였으며, 그 중 결정계수(R2)값이 가장 높은 모델을 선정하였다. 그래프는 SigmaPlot(Sigma Plot 12.5, Systat Software Inc., USA)프로그램을 이용하여 표현하였다. 결정계수(R2)와 평균 제곱근 오차(RMSE)를 기준으로 모델의 정확도를 평가하였다.
결과 및 고찰
정식 이후 20DAT부터 생장이 활발히 이루어졌으며, 27DAT의 작물 초장과 LAI는 각각 81.29cm, 1.662였으며, 62DAT에는 309.32cm, 2.620였다. 또한 85DAT에 작물의 초장과 LAI는 각각 350.53cm와 2.81이었다. LAI는 40DAT를 전후로 적엽 처리에 따라 값의 변동 폭이 감소하였다(Fig. 3).

Fig. 3.
Plant height (A) and leaf area index (B) changes of plants during the experimental period.
생육단계별 정확한 흡광계수 측정을 위하여 재배기간 초기(1-26DAT), 중기(27-63DAT), 말기(64-85DAT) 중에서 일 누적광량이 가장 많은 날을 선정한 후 분석하였다. 생육 초기, 중기, 말기에서 최상단 광량과 최하단의 광량의 차이는 각각 69%, 72%, 81%로 생육단계가 지남에 따라 차이가 증가하는 경향을 보였다. 이는 작물 생장에 따른 엽면적의 증가로 인해 상단부에서 하단부로 갈수록 잎에 의한 차광이 된 것으로 판단되며, 군락 내 주변 작물의 그림자도 영향을 주었을 것으로 사료된다(Fig. 4). Kendrick과 Kronenberg(1994) 및 Monsi와 Saeki(2005)는 작물의 생장 형태에 따른 흡광비율 감소에 대한 영향을 받아, 캐노피 상부에서 하부로 내려갈수록 광량이 기하급수적으로 감소한다고 보고하였다. 본 실험의 결과에서도 작물이 생장함에 따라 하단에 도달하는 광량이 감소하였으며, 특히 60DAT 이후 최하단의 급격한 광량의 감소는 엽간 상호간섭뿐만 아니라 군락내 다른 개체의 그림자가 차광에 영향을 미쳤을 것으로 사료된다. 또한 하루 중에도 높이에 따른 일사량의 증감이 다르게 변화하는 것을 확인할 수 있었으며(Fig. 4), 이는 온실 구조물에 따른 광환경이 영향을 준 것으로 판단되었다.
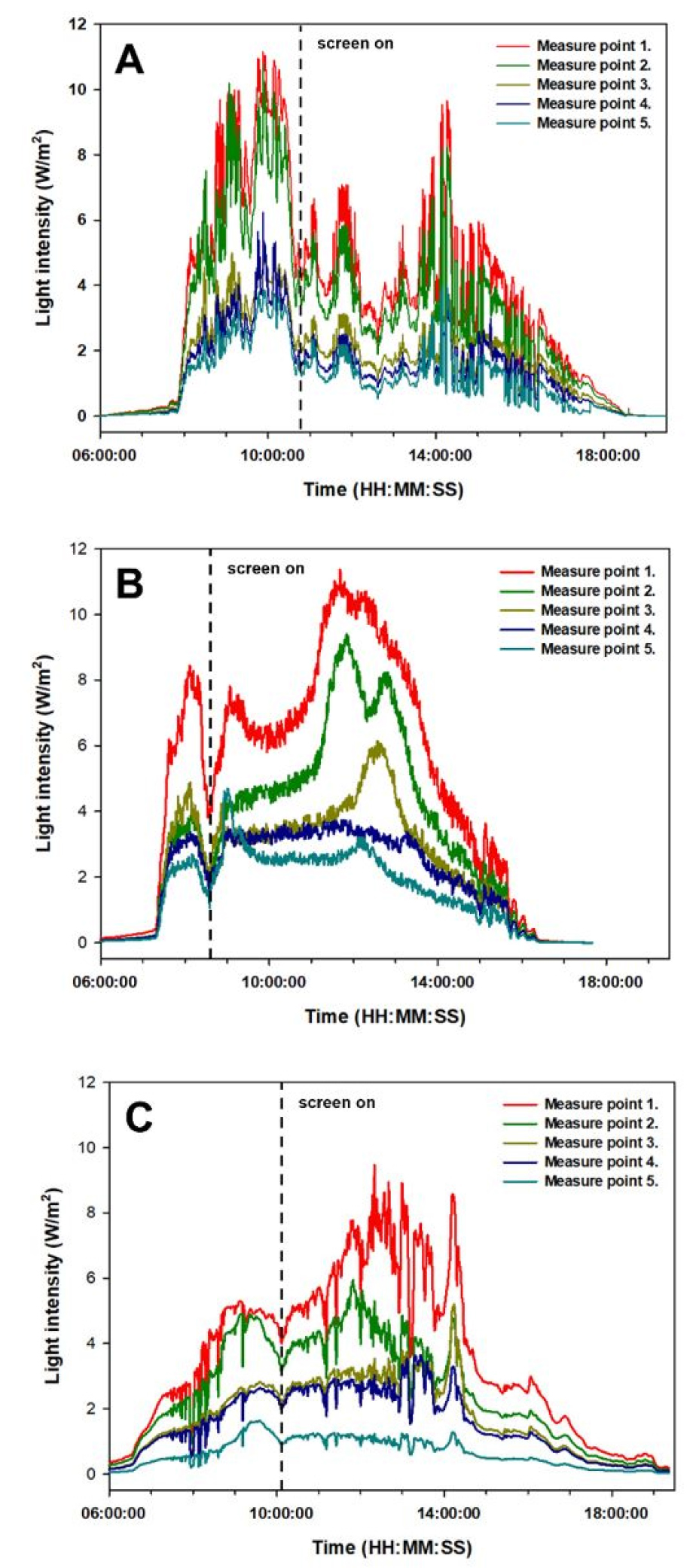
Fig. 4.
Changes of light intensity during the day on April 26, May 17, and June 6 on 2022.
작물 재배일수에 따른 흡광계수 변화는 1-35DAT 구간에서 급격하게 감소하는 경향을 보였고, 40-85DAT 사이 구간의 흡광계수는 큰 폭의 변동을 보이지 않았다(Fig. 5). 1-35DAT의 급격한 흡광계수 감소는 작물의 초기 활착이후 신엽의 활발한 전개가 엽각과 잎의 투과율을 변화시켜 흡광계수에 영향을 미친 것으로 판단된다. Turton(1985), Smith 등(1991), Dufrene와 Breda(1995), Hara(1986), 그리고 Tahiri 등(2006)는 작물의 높이와 재식밀도에 따라 흡광계수가 변화할 수 있다고 보고하였다. 본 실험에서는 작물의 적엽처리 후 엽면적지수의 변화가 거의 없는 40DAT 이후에서 흡광계수의 변화는 군락내 개체들 간의 상호간섭에 의한 것으로 사료되며, 이를 통해 재식밀도에 의한 흡광계수의 변화 또한 고려해야 될 사항으로 판단하였다.
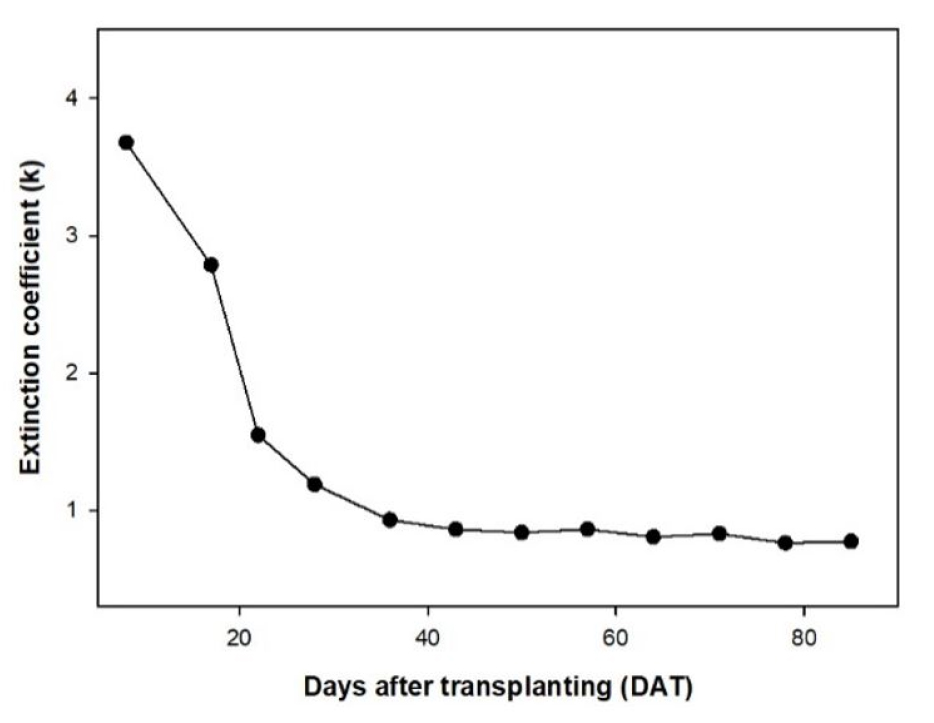
Fig. 5.
Changes of light extinction coefficient according to days after transplanting.
실험기간 동안 작물 LAI와 흡광계수의 관계는 LAI가 0.462일때 k값이 3.678로 가장 높았으며, LAI가 2.621일때는 k값이 0.773로 가장 낮았다(Fig. 6A). 작물의 초장과 흡광계수 관계는 23.49-96.74cm 구간까지는 흡광계수가 급격하게 감소하는 경향을 보였으며, 이후 구간에서는 큰 차이를 보이지 않았다(Fig. 6B). LAI와 초장 두 그래프 모두 지수적 감소(exponential decay) 형태를 나타냈으며 결정계수(R2)는 각각 0.933, 0.939 이었다. 작물의 생장에 따른 LAI와 초장의 증가는 식물 간 상호간섭과, 잎의 방위각 분포를 변화시켜 군락 내 광환경에 영향을 준다고 보고되었다(Loomis 등, 1968; Pepper 등, 1977). 따라서 본 실험의 결과로 미루어 LAI와 초장의 변화가 줄어드는 55DAT를 기준으로 오이의 흡광계수가 수광 특성 파악을 위한 지표 데이터로 이용될 수 있을 것으로 사료된다.

Fig. 6.
Changes of light extinction coefficient according to leaf area index (LAI) and plant height.
LAI와 초장이 증가함에 따라 흡광계수는 감소하는 경향을 보였다(Fig. 7). LAI와 초장의 변화에 따른 흡광계수의 관계는 Paraboloid 식에서 결정계수(R2)와 RMSE 각각 0.968, 1.340으로 관계를 가장 잘 나타냈다(Table 1). Iwakiri와 Inayama(1974), Acock 등(1978), 그리고 Uchijima 등(1976)은 초기에 작물의 잎이 같은 모양과 각도를 가지고 있다고 가정했지만, 토마토, 오이와 같은 작물에서 엽각이 달라 흡광계수의 차이가 생긴다고 하였으며, 또한 온실의 피복재에 의한 확산 광에 의한 변화와 계절에 의한 차이를 보고하였다. 추정식을 통한 예측 값과 실측 값 간의 평균오차는 50DAT 이전에는 22%였지만, 50DAT 이후에는 10%로 이하로 줄어 들었다. 이는 작물 생육초기에 완전히 전개되지 않은 잎에 의한 영향을 완전히 반영하지 못하였지만, 생육 중기 이후에는 작물의 수광형태를 잘 반영하여 광 환경을 잘 추정한 것으로 판단된다.

Fig. 7.
The relationship among the light extinction coefficient, plant height, and leaf area index (LAI).
Table 1.
The regression equation and constant for the relationship among light extinction coefficient (k), plant height (Height), and leaf area index (LAI).
|
Function
|
Regression Equation
|
R2 |
RMSE
|
|
Paraboloid
| |
0.968
|
1.340
|
|
Plane
| |
0.842
|
1.800
|
|
Gaussian
| |
0.951
|
4.180
|
|
Lorentzian
| |
0.902
|
4.599
|
결 론
구해진 추정식을 활용하여 생육단계에 따라 변화하는 일별 흡광계수를 추정할 수 있을 것으로 사료된다. 이를 통해 작물 재배기간 동안 정확한 흡광계수의 예측은 작물의 높이에 따른 광합성 및 증산량 예측 분석에 활용될 수 있을 것으로 판단된다. 또한 작물의 관리방법에 따라 변화하는 광환경과 재식밀도에 따른 흡광계수가 달라질 수 있으므로 이를 고려한 보정방법에 대한 연구가 추가적으로 필요하다.
Acknowledgements
본 연구는 한국스마트팜 R&D재단(과제번호: 421001-03)의 지원에 의해 이루어진 것임.
References
Acock B., D.A. Charles-Edwards, D.J. Fitter, D.W. Hand, L.J. Ludwig, J. Warren Wilson, and A.C. Withers 1978, The contribution of leaves from different levels within a tomato crop to canopy net photosynthesis: an experimental examination of two canopy models. J Exp Bot 29:815-827.
10.1093/jxb/29.4.815Chen J.M., J. Liu, J. Cihlar, and M.L. Goulden 1999, Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecol Model 124:99-119. doi:10.1016/S0304-3800(99)00156-8
10.1016/S0304-3800(99)00156-8Dufrene E., and N. Breda 1995, Estimation of deciduous forest leaf area index using direct and indirect methods. Oecologia 104:156-162.
10.1007/BF0032858028307352Hara T. 1986, Effects of density and extinction coefficient on size variability in plant populations. Ann Bot 57:885-892.
10.1093/oxfordjournals.aob.a087173Higashide T. 2022, Review of dry matter production and growth modelling to improve the yield of greenhouse tomatoes. Hortic J 91:247-266. doi:10.2503/hortj.UTD-R019
10.2503/hortj.UTD-R019Hirose T. 2005, Development of the Monsi-Saeki theory on canopy structure and function. Ann Bot 95:483-494. doi:10.1093/aob/mci047
10.1093/aob/mci04715585544PMC4246794Iwakiri S., and M. Inayama 1974, Studies on the canopy photosynthesis of the horticultural crops in controlled environment: (2) Distribution of percent sunlit leaf area in hedgerow cucumber canopies. J Agric Meteorol 30:17-26.
10.2480/agrmet.30.17Jones H.G. 1992, Plants and microclimate: a quantitative approach to environmental plant physiology, Ed 2. Cambridge University Press, Cambridge, UK, pp 31-35.
Kendrick R.E., and G.H.M Kronenberg 1994, Photomorphogenesis in Plants, Ed 2. Kluwer Academic Publishers, Dordrecht, the Netherlands, pp 99-114.
Loomis R.S., W.A. Williams, W.G. Duncan, A. Dovrat, and F.A. Nunez 1968, Quantitative descriptions of foliage display and light absorption in field communities of corn plants. Crop Sci 8:352-356. doi:10.2135/cropsci1968.0011183X000800030027x
10.2135/cropsci1968.0011183X000800030027xMonsi M., and T. Saeki 2005, On the factor light in plant communities and its importance for matter production. Ann Bot 95:549-567. doi:10.1093/aob/mci052
10.1093/aob/mci05215661751PMC4246799Nederhoff E.M. 1983, Light interception of a cucumber crop at different stages of growth. Acta Hortic 148:525-534. doi:10.17660/ActaHortic.1984.148.66
10.17660/ActaHortic.1984.148.66Nobel P.S., I.N. Forseth, and S.P. Long 1993, Canopy structure and light interception. Photosynthesis and production in a changing environment. A Field and Laboratory Manual. Springer, Dordrecht, the Netherlands, pp 79-90.
10.1007/978-94-010-9626-3_6Peil R.M., M.M. González-Real, and J. López-Gálvez 2002, Light interception of a greenhouse cucumber crop: Measurements and modeling results. Acta Hortic 588:81-87. doi:10.17660/ActaHortic.2002.588.11
10.17660/ActaHortic.2002.588.11Pepper G.E., R.B. Pearce, and J.J. Mock 1977, Leaf orientation and yield of maize. Crop Sci 17:883-886. doi:10.2135/cropsci1977.0011183X001700060017x
10.2135/cropsci1977.0011183X001700060017xRural Development Administration (RDA) 2018, Cucumber-Agricultural technology guide 107. RDA, Korea, pp 24-25.
Smith F.W., D.A. Sampson, and J.N. Long 1991, Comparison of leaf area index estimates from tree allometrics and measured light interception. For Sci 37:1682-1688. doi:10.1093/forestscience/37.6.1682
10.1093/forestscience/37.6.1682Tahiri A.Z., H. Anyoji, and H. Yasuda 2006, Fixed and variable light extinction coefficients for estimating plant transpiration and soil evaporation under irrigated maize. Agric Water Manag 84:186-192. doi:10.1016/j.agwat.2006.02.002
10.1016/j.agwat.2006.02.002Tan C.W., P.P. Zhang, X.X. Zhou, Z.X. Wang, Z.Q. Xu, W. Mao, W.X. Li, Z.Y. Huo, W.S. Guo, and F. Yun 2020, Quantitative monitoring of leaf area index in wheat of different plant types by integrating NDVI and Beer-Lambert law. Sci Rep 10:929. doi:10.1038/s41598-020-57750-z
10.1038/s41598-020-57750-z31969589PMC6976636Turton S.M. 1985, The relative distribution of photosynthetically active radiation within four tree canopies, Cragieburn Range, New Zealand. Aust For Res 15:383-394.
Uchijima Z., K. Inoue, and S. Kimura 1976, The climate in growth chamber: (6) Diffuse radiation environment in vinylhouses. J Agric Meteorol 32:117-125.
10.2480/agrmet.32.117Vos J., J.B. Evers, G.H. Buck-Sorlin, B. Andrieu, M. Chelle, and P.H.B. de Visser 2010, Functional-structural plant modelling: A new versatile tool in crop science. J Exp Bot 61:2101-2115. doi:10.1093/jxb/erp345
10.1093/jxb/erp34519995824

