

서 론
딸기(Fragaria × ananassa Duch.)는 겨울을 대표하는 한국의 시설원예작물 중 하나로 당도가 높고 비타민과 항암물질 등 기능성 물질이 풍부하며 향이 좋아 소비자들의 수요가 높은 작물 중 하나이다(Méndez-Lagunas 등, 2017; Bae 등, 2019). 한국산 딸기 품종과 생과는 특유의 향미와 식미를 해외에서도 인정받아 국내뿐만 아니라 해외 시장에도 많은 양이 수출되고 있다. 2018년도 홍콩과 싱가포르 지역에서는 한국 딸기가 전체 수출 딸기 시장의 63% 비중을 차지하였고(Park 등, 2019), 홍콩, 싱가포르, 베트남, 태국 등의 동남아 시장을 중심으로 2018년도에는 수출량 4,151톤, 수출액 42,792천불, 2019년도에는 수출량 5,033톤, 수출액 49,345천불의 딸기가 수출되어 그 수출량이 점점 증가하고 있음이 확인되었다(KATI, 2020). 현재 한국의 딸기 농가에서는 ‘설향’, ‘죽향’, ‘장희’, ‘매향’ 등 약 7개 품종이 주로 재배되고 있으며(Park 등, 2019), 이 중 ‘매향’은 맛과 향이 좋고 과실의 경도가 높아 저장 및 유통에 유리한 특징을 가지고 있어 전체 수출 품종 중 90% 이상의 비중을 차지하고 있다(Kim 등, 2004; Park 등, 2019; Park 등, 2020). 따라서, 증가하는 해외의 딸기 수요를 충당할 수 있는 생산 능력의 확보가 필요하며, 이를 위해 과실을 생산할 수 있는 ‘매향’ 딸기 자묘의 대량 생산 기술 개발은 중요한 과제라고 할 수 있다.
농가에서 재배되는 딸기는 서로 다른 두 종을 교배하여 만들어진 8배체의 복잡한 유전자형을 가지고 있어(Husaini와 Neri, 2016), 일반적인 과채류처럼 종자번식을 실시할 경우 다음 세대로 품종 특성을 유지하기 어렵고, 종자 발아율 또한 60% 전후로 낮아 경제성이 떨어지는 단점을 갖고 있다(Jurik, 1985; Rugienius와 Stanys, 2001). 이러한 이유로, 일반 농가에서는 딸기 모주에서 발생한 런너를 영양번식하여 자묘를 생산하고 있다(RDA, 2009). 최근 딸기 농가에서는 토양병과 역병을 방지하며, 건전한 묘를 생산하기 위해 과거 노지 육묘 방식이 아닌 비가림 시설에서 육묘를 하는 비가림 포트 육묘 방식으로 자묘를 생산하고 있다(Kim 등, 2012; Kim 등, 2013). 비가림 포트 육묘 방식은 크게 유인 방법과 삽목 방법으로 나누어진다. 유인 방법은 모주와 런너 간 연결을 유지하여 자묘의 생존율을 높일 수 있으나 10월부터 이듬해 9월까지 11개월 이상의 긴 육묘 기간이 소요되고 노동력이 많이 드는 단점이 있다(Hwang 등, 2020b). 반면에, 삽목 육묘 방식은 모주에서 분리한 런너를 삽수로 사용하기 때문에 모주를 관리하는 기간이 없어 육묘 기간이 짧고 노동력이 적게 드는 장점이 있으나 추가적인 가습 시설이 필요하고 삽목 후 수분 관리를 못할 경우 묘가 고사할 위험이 있다(Kang 등, 2019; Hwang 등, 2020b).
일반적으로 삽목번식에서 발근율과 뿌리 발달에 영향을 미치는 요인은 상대습도 유지, 관수 방법, 상토 종류, 그리고 발근제 처리 등이 있는데(Kim과 Kim, 2012), 딸기는 품종에 따른 발근 및 생육의 차이가 크기 때문에(Kim 등, 1999), 이를 고려한 품종 간 적정 육묘 환경 연구가 필요하다. 최근 수출 딸기 품종인 ‘매향’의 경우 삽목 시 가습과 차광처리, 관수 방법, 가습기간과 배지 종류에 관한 연구가 진행되었지만(Kim 등, 2018; Kang 등, 2019; Hwang 등, 2020b), 삽목 시 발근율을 높이기 위한 발근제 처리에 따른 ‘매향’ 품종의 발근 및 생육에 대해서는 아직 보고된 바 없다.
옥신은 식물의 발근제로 많이 상용되는 대표적인 식물생장조절물질 중 하나이며 식물의 세포 신장, 발아, 기관의 분화, 그리고 개화 등에 관여하며 식물 뿌리에 공급 시 뿌리 발근 및 식물 생장을 촉진하는 것으로 알려져 있다(Devender와 David, 1984; Jung 등, 2006). 그중에서도 1-naphthylacetamide(NAD), 3-indoleacetic acid(IAA), 그리고 indole-3-butyric acid(IBA) 등은 국내에서 상용되는 발근제의 재료로써 많이 사용되고 있다(RDANIAS, 2020). 딸기의 경우 내수용으로 많이 재배되고 있는 ‘설향’ 품종의 삽수 채취시기 및 옥신 처리에 따른 발근 및 생육에 관한 연구가 진행된 바 있으며, 6월에 채취한 삽수에 IAA를 1,000mg·L-1 이하의 농도로 처리 시, 삽목묘의 생존율과 생육이 우수하였다고 보고되었다(Kim 등, 2020).
따라서, 본 연구에서는 수출용으로 많이 재배되고 있는 ‘매향’ 딸기 품종에서 옥신의 종류 및 처리 농도에 따른 발근율, 생육 및 뿌리 형태 특성을 조사하고 이를 통해 옥신의 딸기 삽목용 발근제로써 사용 가능성을 확인하기 위해 수행되었다.
재료 및 방법
1. 실험재료 및 재배환경
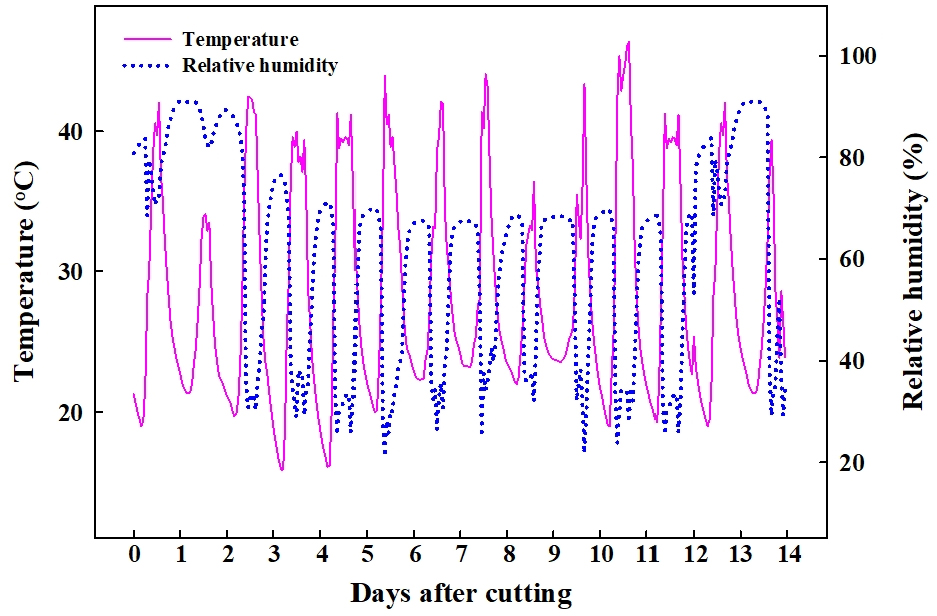
2020년 5월 31일에 ‘매향’ 품종의 모주에서 삽수를 채묘하여 일시 삽목과 화아분화 유도를 위해 5℃ 농업용 저온 냉장고에 4일간 저온 보관한 뒤 2020년 6월 4일에 농도 50, 100, 150, 200mg·L-1로 증류수에 용해한 NAD(1-naphthylacetamide, Sigma-Aldrich Co. Ltd., St. Louis, MO, USA), IBA(indole- 3-butyric acid, Sigma-Aldrich Co. Ltd., St. Louis, MO, USA), 그리고 IAA(3-indoleacetic acid, Sigma-Aldrich Co. Ltd., St. Louis, MO, USA) 수용액에 각각 1시간 동안 침지 처리하였으며, 무처리구를 대조구로 설정하였다. 처리가 끝난 삽수는 48시간 동안 수돗물로 포화시킨 코이어(Coir; Cocopeat Co. Ltd., Dummalasuriya, Sri Lanka)가 충진된 24구 딸기 육묘용 트레이(Hwasung industries Co. Ltd., Okcheon, Korea)에 삽목하였다. 이후 산업용 가습기(UH-303, JB Natural Co. Ltd., Gunpo, Korea)를 설치한 소형 삽목 터널에 14일간 가습 처리를 하였다. 실험기간 동안 온습도 데이터 로거(TR-74Ui, T&D Co. Ltd., Matsumoto, Japan)를 이용하여 소형 삽목 터널 내의 온습도를 측정하였으며, 측정한 소형 삽목 터널 내부의 평균 상대습도는 62.4 ± 15%, 주간 평균온도는 29.3 ± 5℃, 야간 평균온도는 16.2 ± 5℃였다(Fig. 1).
2. 조사항목
삽목 후 14일째에 각 처리 별 삽목묘를 채취하여 발근율, 생존율, 주근 수, 엽병장, 엽장, 엽폭, 엽수, SPAD, 엽면적, 관부 직경, 그리고 지상부와 지하부의 생체중 및 건물중을 측정하였다. 발근율을 비교하기 위해 2mm 이상 자란 새 뿌리만을 발근이 된 것으로 간주하였다. SPAD값은 엽록소 측정기(SPAD- 502, Konica Minolta Inc., Tokyo, Japan)를 이용하여 생장점을 기준으로 완전히 전개된 세 번째 잎을 측정하였다. 엽면적은 엽면적 측정기(LI-3000, LICOR Inc., Lincoln, NE, USA)를 이용하여 측정하였다. 관부 직경을 측정하기 위해 버니어캘리퍼스(CD-20CPX, Mitutoyo Co. Ltd., Kawasaki, Japan)를 사용하였으며, 생체중과 건물중은 전자저울(EW220-3NM, Kern&Sohn GmbH., Balingen, Germany)을 이용하여 측정하였다. 건물중은 시료를 항온 건조기(Venticell-222, MMM Medcenter Einrichtungen GmbH., Planegg, Germany)를 이용하여 70℃에서 72시간 건조 시킨 후 측정하였다. 옥신의 종류와 농도에 따른 삽목묘의 생존율과 뿌리 발달 정도를 확인하기 위해, 삽목 후 21일째에 각 처리별 삽목묘의 생존율을 확인하였으며, 삽목묘의 지하부를 채취하여, WinRhizo Pro 2007a 형태 분석 시스템(Regent Instruments Inc., Sainte-Foy, QC, Canada)을 이용하여 처리별 총 뿌리 길이, 뿌리 표면적, 근단 수, 및 뿌리 분기 수를 측정하였다.
3. 실험설계 및 통계분석
실험은 처리당 48개체씩 3반복의 난괴법으로 배치하였으며 균일한 12개체의 삽목묘를 채취하여 조사하였다. 통계분석은 SAS 프로그램(SAS 9.4, SAS Institute Inc., Cary, NC, USA)을 이용하여 분산분석(ANOVA)을 실시하였고, 평균간 비교는 Tukey 다중검정을 이용하여 5% 유의수준에서 각 처리 간 유의성을 검증하였다. 그래프는 SigmaPlot 프로그램(SigmaPlot 12.5, Systat Software Inc., San Jose, CA, USA)을 이용하여 나타냈다.
결과 및 고찰
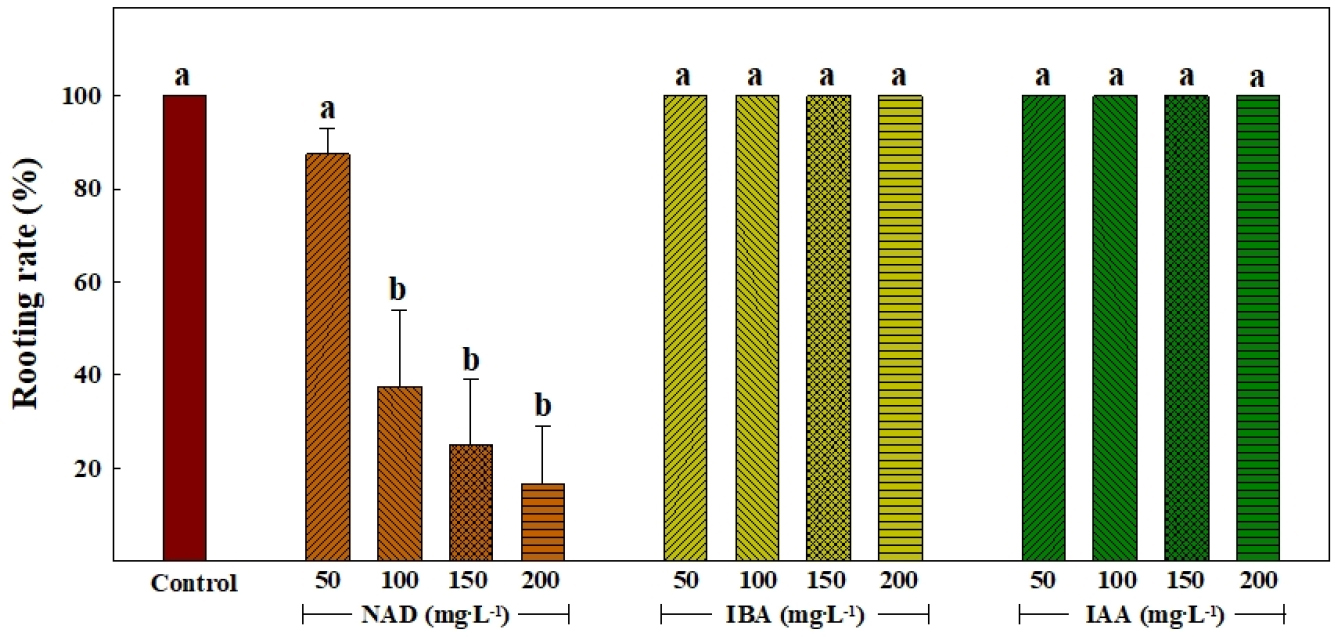
Fig. 2는 삽목 후 14일째 옥신의 종류 및 처리 농도에 따른 삽목묘의 발근율을 조사한 것이다. 대조구와 IBA 및 IAA 처리에서는 모두 100%의 발근율을 나타내었으나, NAD 처리에서 다른 처리에 비해 낮은 발근율을 나타내었으며, 처리 농도가 높아질수록 발근율이 더욱 낮아지는 경향을 보였다. 고농도의 옥신은 세포 수용체의 작용 부위에 비효율적으로 부착되어 세포가 옥신을 정상적으로 수용했을 때 나타나는 세포의 대사 작용 및 생육을 억제하는 경향이 있다(Gardner 등, 2020). 특히 NAA(1-naphthaleneacetic acid)와 같이 나프탈렌 고리를 바탕으로 합성된 NAD는 고리 구조의 차이에 의해 세포막에서의 작용 능력이 떨어진다(Flasiński와 Hąc-Wydro, 2014). 이러한 나프탈렌 고리 바탕의 합성 옥신 처리에 의한 뿌리 발근 저하는 ‘설향’ 딸기묘의 발근 실험에서도 보고된 바 있다(Kim 등, 2020). 따라서, 본 현상은 상대적으로 세포막에서의 작용 능력이 떨어지는 NAD가 고농도로 처리됨에 따라 옥신 대사 장애가 발생하여 정상적인 딸기 묘의 발근이 억제된 것으로 판단된다.
삽목 후 21일째 옥신의 종류 및 처리 농도에 따른 삽목묘의 생존율을 조사하였다(Fig. 3). 대조구는 100%의 생존율을 보였다. IBA 처리는 50mg·L-1 농도에서 다른 농도에 비해 유의적으로 높은 95.8%의 생존율을 보였으며, IAA 처리는 150mg·L-1 이상의 농도를 처리했을 때 생존율이 50% 이하로 낮아지는 것을 확인하였다. IBA 처리에서 IAA 처리보다 150mg·L-1 이상의 고농도에서 안정적인 생존율을 보이는 것을 확인할 수 있다. IBA는 천연옥신(IAA)의 전구물질로서 식물에 흡수되며(Van der Krieken 등, 1993), 식물체 내에서 천연옥신과 별개의 이동 및 작용기작을 거치기 때문에(Strader와 Bartel, 2011), 농도에 따른 식물체 내 천연옥신의 이동 및 대사에 비교적 끼치는 영향이 적다(Strader 등, 2011; Strader와 Bartel, 2011). 그러므로, 세포에 흡수되어 바로 옥신 대사에 관여하는 IAA와 다르게 IBA는 고농도에서도 안정적인 생존율을 보이는 것으로 판단된다. NAD 처리는 전체적으로 가장 낮은 생존율을 보였고 처리 농도가 100mg·L-1 이상일 때 생존율은 0%였다. 이는 앞서 조사된 낮은 발근율과 이에 따른 늦은 착근에 의한 결과로 여겨진다.
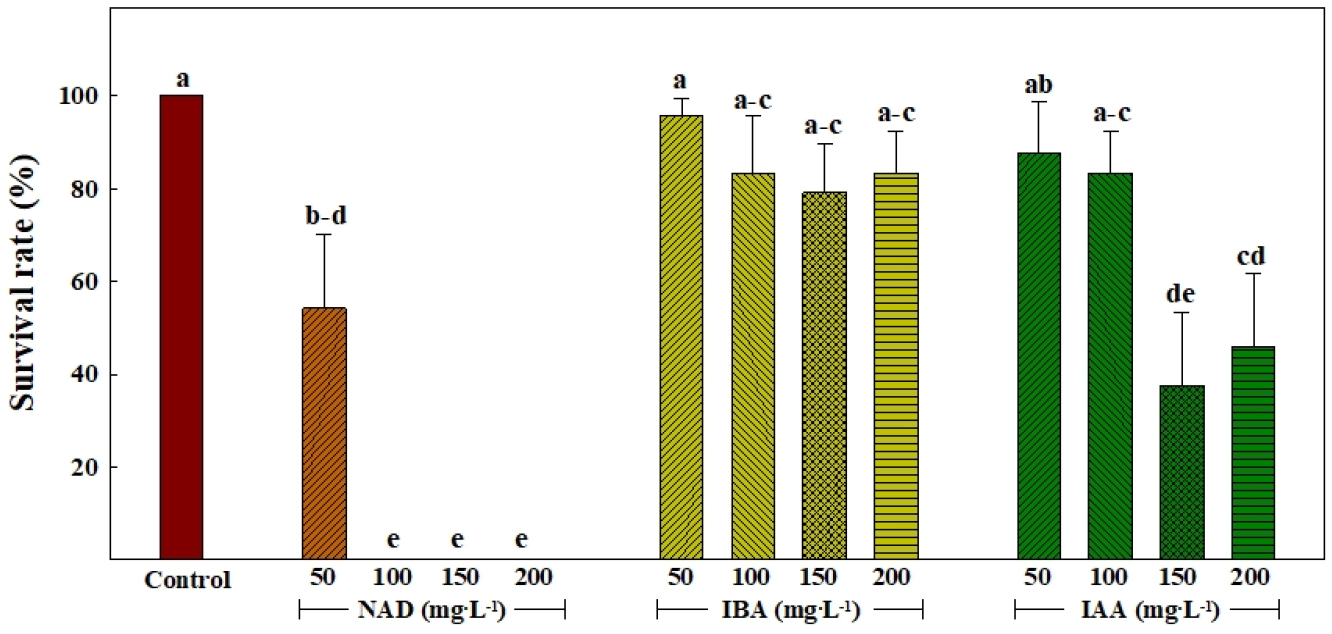
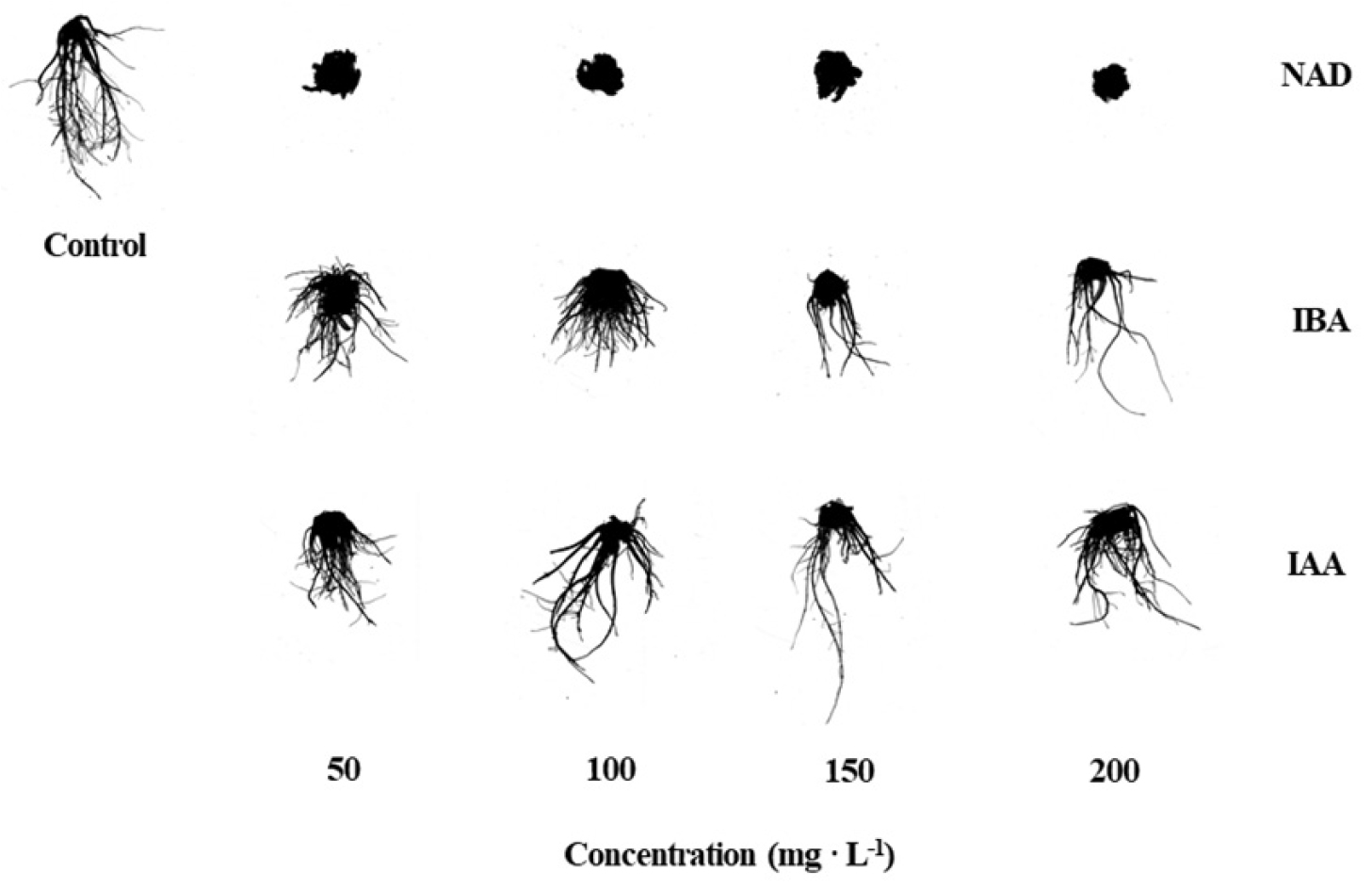
Fig. 4는 삽목 후 14일째 옥신의 종류 및 처리 농도에 따른 삽목묘의 발근 및 생육 상태를 나타낸 것이며, 삽목 후 14일째 옥신의 종류 및 처리 농도에 따른 삽목묘의 지상부 생육을 Table 1에 나타내었다. 엽병장, 엽장, 엽폭, 관부직경 및 SPAD는 처리 간 유의적인 차이가 없었다. Hwang 등(2020a)은 ‘매향’ 품종에 IBA를 처리했을 때 처리 간 지상부 생육 및 런너 신장에서 유의적인 차이가 없었다고 보고하였으며, Kim 등(2020)도 ‘설향’ 딸기에 옥신을 처리하였을 때 지상부 생육에서 유의적인 차이가 없었다고 보고하였다. Gardner 등(2020)은 작물의 품종과 기관별 옥신에 대한 감응성이 달라질 수 있다고 하였으며, Murti와 Yeoung(2013)은 딸기에서 품종에 따른 옥신 처리에 대한 생육 경향성에서 차이가 날 수 있다고 보고하였다. 엽수는 IBA 처리 100mg·L-1 농도에서 가장 많았으며, 100mg·L-1 이상의 농도에서 감소하였다. 옥신은 조직 내 축적 정도에 따라 세포에 영양을 집중시켜 조직의 분화 및 발달을 활발하게 할 수 있으나(Krouk 등, 2010), 최적 농도 이상 처리될 경우 오히려 생육을 억제하게 된다(Gardner 등, 2020). 따라서, IBA 처리 100mg·L-1 농도가 ‘매향’ 딸기의 잎 세포 발달 및 분화에 적절한 농도인 것으로 판단된다. Table 2는 삽목 후 14일째 옥신의 종류 및 처리 농도에 따른 삽목묘의 지상부와 지하부 생체중 및 건물중과 엽면적을 나타낸 것이다. 지하부 건물중은 대조구에서 0.17g으로 유의적으로 가장 무거웠으며, NAD 처리에서 다른 옥신 처리에 비해 농도에 관계없이 평균 0.11g으로 유의적으로 가장 낮은 건물중을 보였다. 지상부의 생체중과 건물중, 지하부 생체중, 그리고 엽면적은 처리 간 유의적인 차이가 없었다. 이는 앞서 옥신 처리에 따른 대부분의 지상부 생육에서 유의적인 차이가 없었기 때문인 것으로 판단된다.

Table 1.
Growth characteristics of cutting propagated ‘Maehyang’ strawberry as affected by various types and concentrations of auxin at 14 days after treatments (n = 12).
|
Treated auxinz (A) |
Concentration (mg·L-1) (B) |
Petiole length (cm) |
Leaf length (cm) |
Leaf width (cm) | No. of leaves |
No. of primary roots |
Crown diameter (mm) | SPAD |
| Control | 11.2 | 5.7 | 4.6 | 3.3 a-cy | 22.3 c-e | 6.8 | 37.2 | |
| NAD | 50 | 12.8 | 6.2 | 4.6 | 2.6 c | 32.4 a-c | 6.1 | 37.9 |
| 100 | 13.7 | 6.1 | 4.7 | 2.8 a-c | 23.0 b-e | 7.3 | 39.0 | |
| 150 | 11.5 | 5.6 | 4.4 | 3.1 bc | 11.5 a-c | 6.4 | 36.7 | |
| 200 | 13.5 | 6.3 | 4.7 | 2.6 c | 13.9 de | 6.7 | 36.4 | |
| IBA | 50 | 12.9 | 6.1 | 5.0 | 3.2 a-c | 21.8 c-e | 7.0 | 34.9 |
| 100 | 12.8 | 5.7 | 4.4 | 3.5 a | 34.1 a-c | 6.7 | 33.4 | |
| 150 | 11.8 | 6.4 | 4.9 | 2.8 a-c | 40.8 ab | 6.5 | 38.7 | |
| 200 | 14.3 | 6.8 | 5.3 | 2.8 bc | 31.2 a-d | 6.0 | 37.2 | |
| IAA | 50 | 12.4 | 6.2 | 4.6 | 3.1 a-c | 23.0 b-e | 5.5 | 39.1 |
| 100 | 13.3 | 5.9 | 4.6 | 2.8 bc | 44.6 a | 5.6 | 37.5 | |
| 150 | 11.9 | 5.4 | 4.4 | 2.9 a-c | 46.1 a | 6.1 | 41.6 | |
| 200 | 13.5 | 6.3 | 5.1 | 3.3 ab | 34.1 a-c | 6.3 | 36.7 | |
| F-test | A | NS | NS | NS | * | *** | NS | NS |
| B | NS | NS | NS | NS | * | NS | NS | |
| A×B | NS | NS | NS | *** | *** | NS | NS |
Table 2.
The fresh and dry weights, and leaf area of cutting propagated ‘Maehyang’ strawberry as affected by various types and concentrations of auxin at 14 days after treatments (n = 12).
| Treated auxinz (A) | Concentration (mg·L-1) (B) | Fresh weight (g/plant) | Dry weight (g/plant) | Leaf area (cm2/plant) | |||
| Shoot | Root | Shoot | Root | ||||
| Control | 3.68 | 1.02 | 0.90 | 0.17 ay | 109.87 | ||
| NAD | 50 | 3.46 | 0.99 | 0.89 | 0.11 b | 112.69 | |
| 100 | 4.25 | 1.07 | 1.01 | 0.12 ab | 116.91 | ||
| 150 | 3.38 | 0.96 | 0.76 | 0.10 b | 79.21 | ||
| 200 | 6.37 | 1.10 | 0.80 | 0.11 b | 94.74 | ||
| IBA | 50 | 3.97 | 0.97 | 0.88 | 0.13 ab | 126.91 | |
| 100 | 3.80 | 1.02 | 0.94 | 0.15 ab | 120.11 | ||
| 150 | 3.92 | 0.96 | 0.86 | 0.13 ab | 108.44 | ||
| 200 | 4.31 | 1.12 | 1.01 | 0.14 ab | 119.64 | ||
| IAA | 50 | 3.41 | 0.94 | 0.84 | 0.14 ab | 107.31 | |
| 100 | 3.42 | 0.86 | 0.80 | 0.13 ab | 113.94 | ||
| 150 | 3.59 | 1.05 | 0.85 | 0.15 ab | 109.33 | ||
| 200 | 4.03 | 1.12 | 0.88 | 0.16 ab | 118.42 | ||
| F-test | A | NS | NS | NS | *** | NS | |
| B | NS | NS | NS | NS | NS | ||
| A×B | NS | NS | NS | NS | NS | ||
Fig. 5는 삽목 후 21일째 옥신의 종류 및 처리 농도에 따른 삽목묘의 뿌리 형태를 측정한 모습이며, Fig. 6은 이를 바탕으로 뿌리 형태 분석을 실시한 것이다. 총 뿌리 길이, 근단 수, 뿌리 표면적 및 뿌리 분기 수는 대조구에서 모두 유의적으로 가장 높은 값을 나타내었으며, NAD 처리에서 농도에 관계없이 유의적으로 가장 낮은 값을 보였다. 대조구를 제외하고 옥신 처리가 된 ‘매향’ 딸기의 총 뿌리 길이는 IBA와 IAA 처리 100mg·L-1 농도에서 각각 49.4와 49.9mm로 유의적으로 길었으며, 근단 수는 IAA 처리 100mg·L-1 농도에서 67개로 유의적으로 많았고, 뿌리 표면적과 뿌리 분기 수는 IBA 처리 100mg·L-1 농도에서 각각 13.0cm2와 197개로 유의적으로 높은 값을 보였다. 또한 모든 항목에서 IBA와 IAA 처리는 처리 농도가 150mg·L-1 이상일 때 감소하는 경향을 보였다. 옥신은 삽수의 상처 조직의 치유 및 미분화 세포의 발근 유도를 하는 것으로 알려져 있으며, 실제로 이를 위해 농가에서 삽목용 발근제로 많이 사용하는 식물생장조절물질이다(Gardner 등, 2020). 하지만, 미분화 세포의 발근 유도와 달리 옥신 처리는 발근된 뿌리의 신장 및 생육에 있어 동화 산물 분배 정도에 따른 뿌리 생육 억제 현상을 동반할 수 있으며, 특히 최적 농도 이상의 옥신 처리는 뿌리에서의 에틸렌 발생량 또한 증가시켜 뿌리 생육 억제를 심화할 수 있다(Chadwick과 Burg, 1967; Gardner 등, 2020). 딸기는 가지나 잎을 삽수로 사용하는 타 원예작물과 달리, 유근이 발생한 자묘를 삽수로 사용하기 때문에(Saito 등, 2008; Zheng 등, 2019), 옥신의 발근 유도 보다 뿌리 생육 억제 효과가 더 강하게 작용하였을 것으로 판단된다.
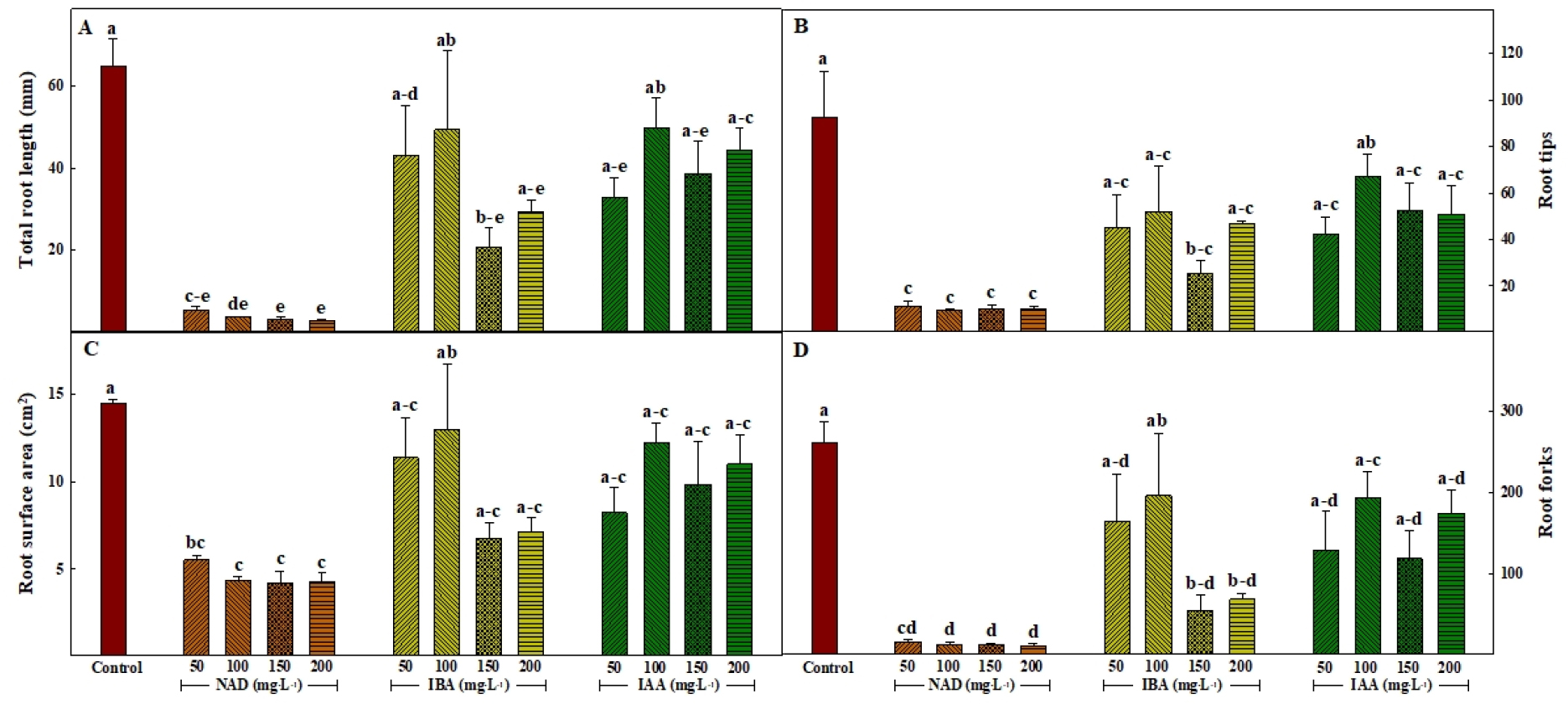
결론적으로 NAD 처리를 한 ‘매향’ 삽목묘는 발근이 제대로 이뤄지지 않았고 삽목묘의 생존율 또한 가장 저조했으며, IAA 처리는 옥신을 처리한 다른 처리들에 비해 많은 주근 수를 보였고, IBA 처리는 높은 생존율과 엽수를 보였다. 대조구는 유의적으로 높은 발근율과 생존율을 보였으며, 총 뿌리 길이, 뿌리 표면적, 근단 수 및 뿌리 분기 수, 그리고 지하부 건물중에서 옥신 처리구에 비해 높은 값을 보여 우수한 뿌리 생육을 확인할 수 있었다. 따라서, 옥신 기반의 딸기 삽목 발근제 개발을 위해서는 NAD보다 IAA나 IBA를 사용하는 것이 좋으며, 유근이 어느 정도 발생한 자묘를 삽수로 이용하는 딸기 삽목법의 특징을 고려하였을 때 단일 옥신 처리가 아닌 다른 생장조절물질을 혼용한 연구가 필요할 것으로 판단된다.
