

서 론
재료 및 방법
1. 실험 재료 및 재배환경
2. 차광 및 보광 처리
3. 생육조사
4. 통계분석
결과 및 고찰
1. 온실 내부의 광도, 온도, 상대습도
2. 오이 생육 특성
3. 오이 과실 특성
서 론
오이(Cucumis sativas L.)는 저온성 작물로 온도와 같은 환경조건에 민감하여 체계적인 관리 기술이 요구되는 과채류 작물이다(RDA, 2018a). 또한, 같은 포기에 암꽃과 수꽃이 분리된 자웅이화이며 저온단일 조건에서 암꽃 분화가 촉진되는 작물로, 암꽃 착생에 유리한 환경조건을 조성하는 것은 중요하다(RDA, 2018a). 주로 오이 과실은 하절기 중에서도 혹서기에 가격이 높게 형성되는데, 하절기는 고온장일 조건의 환경으로 인해 암꽃 착생이 억제되고 생산량이 감소할 수 있다(AT, 2020). 따라서 하절기 냉방 기술을 이용한 온도 하강과 같은 환경조건 개선은 암꽃 착생을 증가시켜 생산량 증대에 도움이 될 것으로 판단된다.
하절기의 온실 내부 온도는 시설과채류의 생육 한계온도인 35℃ 이상으로 지속되기 때문에 고온장해, 수확량 감소 등의 문제를 야기한다(Kim 등, 2020; Lee 등, 2021; Park 등, 2020). 또한, 하절기의 강한 일사량은 오이의 기공을 폐쇄시켜 CO2 흡수를 억제하며, 호흡량을 증가시키기 때문에 광합성 효율이 감소하여 생육 불량, 품질 저하 등의 문제가 발생한다(Cheong 등, 2015; Woo 등, 2014). 따라서 안정적인 하절기 오이 재배를 위해 온실 내부로 유입되는 광량을 감소시키고, 온도를 낮추는 방법을 적용해야 한다. 냉방기 사용은 온실의 온도를 하강시키는 가장 효과적인 방법이지만, 설치 및 운영비용이 과도하게 발생하므로 상대적으로 비용이 적게 발생하는 환기, 차광 그리고 포그냉방 등과 같은 기술이 도입되어 왔다(Kim 등, 2006). 차광은 국내 온실재배에서 가장 많이 활용하고 있는 냉방 방법으로, 온실 내부 온도를 낮춰 개화 및 수정 불량 등의 생육 장해를 감소시켜 작물의 품질과 생산성을 향상시킬 수 있는 장점이 있다(Ha 등, 2012; Lim 등, 2021).
주로 하절기에 발생하는 폭우, 장마 등의 구름이 많은 기상 조건은 온실 내부에 저일조 환경을 유발한다(Lee 등, 2016). 또한, 온실 내부 온도 하강을 위해 설치하는 냉방 장치와 골조구조물로 인해 발생하는 그늘에 의해 유입되는 광량이 감소한다(Jeong 등, 2008; Jeong 등, 2009). 광량 감소는 오이와 같은 시설작물에 광 부족 현상을 유발하여 도장, 낙화를 발생시키고, 생산량이 감소하는 원인이 된다(Woo 등, 2005). 따라서 작물의 광합성과 생육 촉진을 위해 시설의 부족한 광량을 보충해 주는 보광 기술의 적용이 필요하다.
보광은 인공 광원을 이용하여 부족한 광량을 보충하는 기술로, 광원의 종류에 따라 광질, 광량이 다르기 때문에 식물의 생육에 다양한 영향을 미친다(RDA, 2018b). 주로 사용되는 인공 광원은 메탈할라이드등(metal halide lamp), 발광 플라즈마등(lighting emitting plasma), 고압나트륨등(high-pressure sodium lamp)으로, 다양한 광원을 이용한 작물 재배에 관한 연구가 진행되었다(Lee 등, 2014). 고압나트륨등은 광도가 높고, 생육을 촉진하는 효과가 있으나 전력 소모량이 많고, 광합성에 필요한 청색광(400-500nm)의 비율이 낮다는 문제점이 있다(Blom과 Ingratta, 1984; Wheeler 등, 1991). 때문에, 최근에는 발열량이 낮고, 가격이 저렴하며, 수명이 길고, 식물 생육과 형태형성에 필요한 특정 파장만을 선택적으로 조사할 수 있는 발광다이오드(light-emitting diode)를 광원으로 이용하고 있다(An 등, 2011; Hwang 등, 2022).
이전의 과채류 작물 재배를 위한 보광 연구는 약광기인 봄과 겨울철에 보광하는 것에 초점이 맞춰져 있으며, 고온기인 여름철 보광에 대한 연구는 부족한 실정이다. 따라서 본 연구에서는 하절기 오이 온실재배 시 차광과 보광 효과를 구명하고 하절기 보광의 적용 가능성을 확인하였다.
재료 및 방법
1. 실험 재료 및 재배환경
본 실험은 경상국립대학교 부속농장 내 플라스틱 온실에서 2022년 8월 30일부터 2022년 10월 20일까지 51일간 수행하였다. 실험에 사용한 오이 묘는 접수 ‘낙원성청장’(Cucumis sativus L. ‘NakWonSeongcheongjang’, Wonnong Seed Co., Ltd., Anseong, Korea)과 대목 ‘파워 반짝이’(Cucurbita moschata ‘Powerbanjjagi’, Wonnong Seed Co., Ltd., Anseong, Korea)를 접목한 묘를 코이어(Cocopeat Co., Ltd., Dummalasuriya, Sri Lanka)가 충진된 직경 23cm 포트에 1주씩 정식하였다. 정식 후 유인, 적엽, 측지 제거, 병해충 방제 등의 관리 작업은 재배 매뉴얼에 따라 실시하였고(RDA, 2018a), 온실 내 온도가 35℃ 이상일 때는 환기 팬이 자동으로 작동하도록 설정하였다. 과실은 2022년 10월 01일부터 2022년 10월 16일까지 3일 간격으로 총 5회 수확하였다. 재배기간 동안 양·수분 공급은 온실 다용도 액비를 pH 6.0과 EC 1.5dS·m-1로 조제하여 매일 오전 9시, 11시, 오후 1시, 총 3회(회 당 1L씩) 점적관수 하였다(Table 1). 온실 내부의 온·습도와 광도는 실험 기간 동안 매일 1시간 간격으로 측정하였고, Fig. 1의 그래프에 일평균 온·습도와 광도로 나타내었다. 측정 장치는 환경 측정 센서(aM-31, Wise Sensing Inc., Yongin, Korea)를 사용하였다.
Table 1.
Composition of the nutrient solution used in the experiment.

2. 차광 및 보광 처리
차광 처리는 온실 내부 차광 스크린(50%)을 이용하여 플라스틱 온실 1동은 대조구, 1동은 차광 처리구로 설정하였다(Fig. 2). 차광 처리구 온실은 내부의 온도와 상대습도 변화에 관계없이 전일 차광 되도록 설정하였다.

보광 처리는 2022년 9월 7일부터 2022년 10월 20일까지 43일간 실시하였다. 광원 종류에 따라 오이의 생육과 과실에 미치는 영향을 조사하고자 고압나트륨등(HPS, SON-T Agro 400W, Philips, Eindhoven, The Netherlands), White LED(W LED, R:G:B = 5:3:2, PW150-full spectrum, Bissol Co., Ltd., Seoul, Korea), 그리고 Red와 Blue 혼합 LED(RB LED, R:B = 7:3, PW150-targeted spectrum, Bissol Co., Ltd., Seoul, Korea)를 사용하였으며(Fig. 3), 비교를 위해 무처리를 대조구(control)로 설정하였다. 각 광원의 광질은 광파장 측정기(ILT950, International Light Technologies Inc., Peabody, MA, USA)를 이용하여 측정하였다(Fig. 4). 보광을 위한 광도는 식물 생장점 위치에서 150±20μmol·m-2·s-1가 되도록 설정하였다. 보광 시간은 일출 전과 일몰 후 각각 2시간씩, 총 4시간 동안 실시하였으며, 일출과 일몰 시각은 천문우주지식 정보를 기준으로 광원이 점등되도록 설정하였다(KASI, 2022).


3. 생육조사
차광과 보광 처리에 따른 오이의 생육을 비교하기 위해 초장, 엽장, 엽폭, 엽수, 마디수, 경경, 그리고 SPAD를 측정하였다. 초장은 식물체의 지제부에서 생장점까지의 길이를, 엽장과 엽폭은 가장 큰 잎에서 측정하였다. 경경은 지제부로부터 1cm 떨어진 위치에서 버니어캘리퍼스(CD-20CPX, Mitutoyo Co., Ltd., Kawasaki, Japan)를 이용하여 측정하였다. SPAD 값은 엽록소 측정기(SPAD-502, Konica Minolta Inc., Tokyo, Japan)를 이용하여 완전히 전개된 3개의 잎에서 측정하여 평균값을 산출하였다.
오이의 과실 특성을 조사하기 위해 과장, 과경, 과실의 생체중 및 건물중을 측정하였다. 과실은 상품과 기준인 과장 20cm 이상의 과실을 수확하여 조사하였다(RDA, 2018a). 과실 수확 기간 동안 측정한 과실 무게와 개수를 기반으로 평균 과중과 누적 과실 개수도 기록하였다. 과경은 버니어캘리퍼스(CD-20CPX, Mitutoyo Co., Ltd., Kawasaki, Japan)를 이용하여 과실의 가장 굵은 부분을, 과실의 생체중과 건물중은 전자저울(EW220-3NM, Kern & Sohn GmbH., Balingen, Germany)을 이용하여 측정하였다. 건물중은 70℃ 항온 건조기(Venticell-222, MMM Medcenter Einrichtungen GmbH., Planegg, Germany)에서 72시간 건조한 후 측정하였다.
4. 통계분석
통계분석은 SAS 프로그램(SAS 9.4, SAS Institute Inc., Cary, NC, USA)을 사용하여 수행하였다. 실험 결과는 분산 분석(ANOVA)을 실시하였고, F-test를 이용하여 차광과 보광 처리 간 유의성을 검증하였다. 평균 간 비교는 Tukey 다중검정(Tukey’s test)과 최소유의차 검정(least significant difference)을 이용하였다. 그래프는 SigmaPlot 프로그램(SigmaPlot 14.5, Systat Software Inc., CA, USA)을 이용하여 나타냈다.
결과 및 고찰
1. 온실 내부의 광도, 온도, 상대습도
실험기간 동안 차광 처리에 따른 온실 내부의 광도와 온도, 상대습도를 측정하였다(Fig. 1). 차광 처리 온실은 차광 무처리 온실과 비교하여 온실 내부로 유입되는 광량이 약 50% 정도 감소하였다. 차광 50% 온실은 0% 온실과 비교하여 약 1-2℃ 정도 더 낮았고, 상대습도는 차광 0% 온실에서 평균 약 61%, 차광 50%에서 평균 약 62%로 처리구간의 차이가 없었다. 차광은 온실 내부로의 광 유입을 감소시키는 원리를 이용하여 온도를 조절하는 방법이다(Chris, 2011). 특히, 열을 방출하는 적외선의 유입을 효과적으로 감소시켜 온실의 내부 온도를 낮출 수 있다(Ha 등, 2012). 본 연구의 결과에서도 차광 처리에 의해 온실 내부의 온도가 감소하는 것을 확인하였다.
2. 오이 생육 특성
보광 처리 35일 후 오이의 초장, 엽장, 엽폭은 차광 처리구에서 유의성 있게 증가하였고, 엽수는 유의적인 차이가 없었으며, SPAD 값은 RB LED를 제외하고 차광에 의해 증가하는 경향을 보였다(Table 2). 보광 광원에 있어서는 차광 0%에서 HPS가 초장과 엽장이 가장 작았고, 경경은 차광 0%에서 RB LED가 보광 대조구와 다른 처리구 비해 유의적으로 높은 값을 보였다. 강광 환경은 식물체 내의 옥신의 광분해를 촉진시켜 식물 초장을 감소시키고, 약광 환경은 옥신의 광분해를 억제하여 식물의 초장을 증가시킨다(Gardner 등, 2020). 식물은 광량이 적을 때 순광합성량을 증가시키기 위해 엽면적을 넓혀 잎이 차지하는 비율을 증가시킨다(Lusk, 2002). Ha 등(2012)은 파프리카에 차광 시 엽온 강하로 인한 증산량 감소가 줄기세포 신장을 촉진하여 초장이 증가하였다고 보고하였다. Haque 등(2009)은 차광률이 높아질수록 오이 잎의 SPAD 값과 엽면적이 증가하였다고 보고하였다. 본 연구에서도 차광 처리로 인한 온실 내부의 광량과 온도 감소가 오이의 초장, 엽장, 엽폭, SPAD 값 증가에 영향을 미친 것으로 판단된다. Guo 등(2016)은 장미에 HPS를 이용하여 보광 했을 때 광원으로부터 방출되는 열에 의한 엽온 증가로 잎의 크기가 작아졌다고 보고하였다. Hernández와 Kubota(2014)는 높은 DLI(daily light integral, 일일적산광량) 조건에서 오이 묘에 보광 했을 때 RB LED의 적색광 비율이 높을수록 경경이 유의하게 증가하였다고 보고하였다. 본 연구에서도 오이의 엽장은 HPS를 보광 광원으로 이용하였을 때 가장 작았으며, 경경은 차광 0%에서 W LED보다 적색광 비율이 높은 RB LED에서 경경이 가장 굵었다. Bergstrand와 Schüssler(2013)는 온실에서 보광의 효과는 자연광의 양에 따라 달라지는데, 가을철에 비해 광량이 증가하는 봄철 재배에서 보광의 효과가 없었다고 보고하였다. 본 연구는 자연광이 충분한 늦여름 시기에 진행되었기 때문에 보광 대조구와 처리구간의 유의차가 명확하게 나타나지 않은 것으로 판단된다.
Table 2.
Growth characteristics of cucumber as affected by shading level and different supplemental lighting sources at 35 days after supplemental lighting (n = 6).
|
Shading level (A) | Light sources (B) |
Plant height (cm) |
Leaf length (cm) |
Leaf width (cm) |
Number of leaves |
Number of nodes |
Stem diameter (mm) | SPAD |
| 0% | Control | 249.5 abz | 22.3 bc | 28.9 b | 23.5 | 22.8 ab | 11.3 ab | 64.2 ab |
| HPS | 190.8 e | 19.3 d | 27.3 b | 25.0 | 24.5 ab | 11.8 ab | 36.6 b | |
| W LED | 216.6 c-e | 20.3 cd | 27.9 b | 25.8 | 25.2 a | 11.6 ab | 53.8 ab | |
| RB LED | 198.2 de | 22.0 b-d | 29.7 b | 24.2 | 23.7 ab | 13.6 a | 74.4 a | |
| 50% | Control | 260.9 a | 25.0 a | 34.4 a | 24.2 | 23.2 ab | 10.7 ab | 70.8 a |
| HPS | 225.7 b-d | 24.0 ab | 33.1 a | 24.2 | 23.2 ab | 11.4 ab | 76.9 a | |
| W LED | 253.2 ab | 24.1 ab | 33.5 a | 23.2 | 22.2 b | 10.1 b | 72.3 a | |
| RB LED | 236.6 a-c | 25.7 a | 35.1 a | 24.8 | 23.8 ab | 12.0 ab | 59.9 ab | |
| F-test | A | *** | *** | *** | NS | * | * | ** |
| B | *** | *** | ** | NS | NS | * | NS | |
| A×B | NS | NS | NS | NS | * | NS | *** |
3. 오이 과실 특성
과장, 과경, 과실 생체중과 건물중은 보광 대조구와 처리구 간의 유의적인 차이는 없었다(Table 3). 오이 평균 과중은 차광 0%에서 HPS가 가장 낮았고, 차광 50%에서는 10월 8일 수확일을 제외하고 보광 처리에 따른 유의적인 차이는 없었다(Fig. 5). 식물체 당 누적 과실수는 차광 0%의 보광 대조구에서 평균 6개로 가장 많았으나, W, RB LED 간의 통계적으로 유의적인 차이는 없었으며, 차광 50%의 보광 대조구에서는 평균 3개로 가장 많았고, W LED가 가장 적었다(Fig. 6). 오이는 10-15℃의 저온과 8-10시간 이하의 단일 조건에서 암꽃 발현이 촉진되는 특성을 가진다(RDA, 2018b). 본 실험에서는 일출 전, 일몰 후 2시간씩 총 4시간을 보광 했다. 이로 인해 일장이 길어져 암꽃 분화 지연과 과실 개수에 영향을 미쳤고, 보광 대조구의 과실 개수가 가장 많았던 이유로 판단된다. An 등(2011)은 파프리카에 보광을 하였을 때, 엽온 상승으로 인해 동화 산물 전류가 원활하지 못하여 보광 대조구의 착과수가 많았다고 보고하였다. 오이 과실은 낮보다 밤에 왕성하게 비대하는데, 특히 일몰 후 4-5시간 사이에 광합성 동화산물을 Sink 기관으로 옮기는 전류가 활발해진다(RDA, 2018a). 본 연구에서도 일몰 후 2시간 보광이 과실 비대를 위한 전류 과정에 영향을 주어 보광 대조구의 과실 개수가 더 많았던 것으로 판단된다.
Table 3.
Fruit characteristics of cucumber as affected by shading level and different supplemental lighting sources at 36 days after supplemental lighting.
|
Shading level (A) | Light sources (B) |
Fruit length (cm) |
Fruit diameter (mm) | Fruit | |
| Fresh weight (g) | Dry weight (g) | ||||
| 0% | Control | 27.1 az | 38.7 | 203.7 | 6.3 |
| HPS | 25.1 ab | 45.3 | 151.2 | 4.6 | |
| W-LED | 26.4 ab | 36.2 | 189.5 | 4.6 | |
| RB-LED | 23.1 b | 36.4 | 190.7 | 4.8 | |
| 50% | Control | 26.1 ab | 33.7 | 160.0 | 5.2 |
| HPS | 28.1 a | 32.7 | 157.7 | 5.1 | |
| W LED | 26.2 ab | 34.6 | 134.4 | 4.8 | |
| RB LED | 25.1 ab | 34.3 | 159.5 | 3.8 | |
| F-test | A | NS | NS | NS | NS |
| B | * | NS | NS | NS | |
| A×B | NS | NS | NS | NS | |
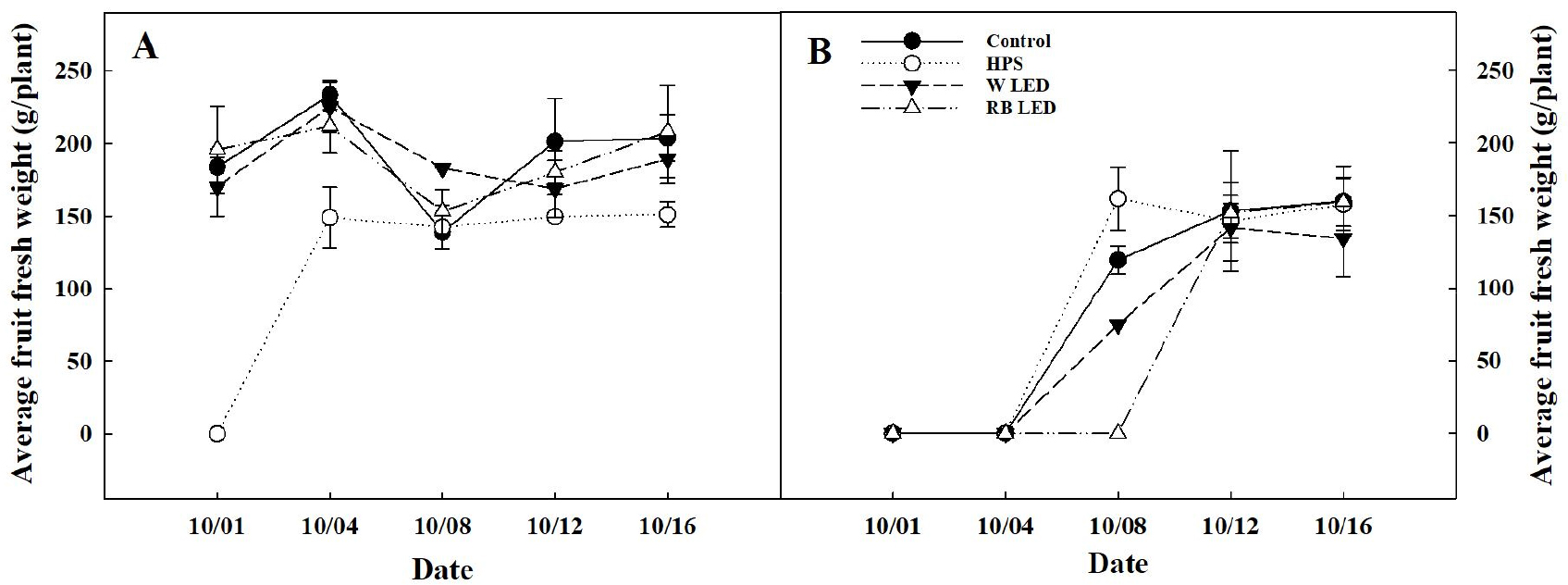
Fig. 5.
Average fruit weight of cucumber as affected by shading level under different supplemental lighting sources. A, 0% shading level and B, 50% shading level. Refer to Fig. 4 for details on spectral distribution of supplemental lighting sources.
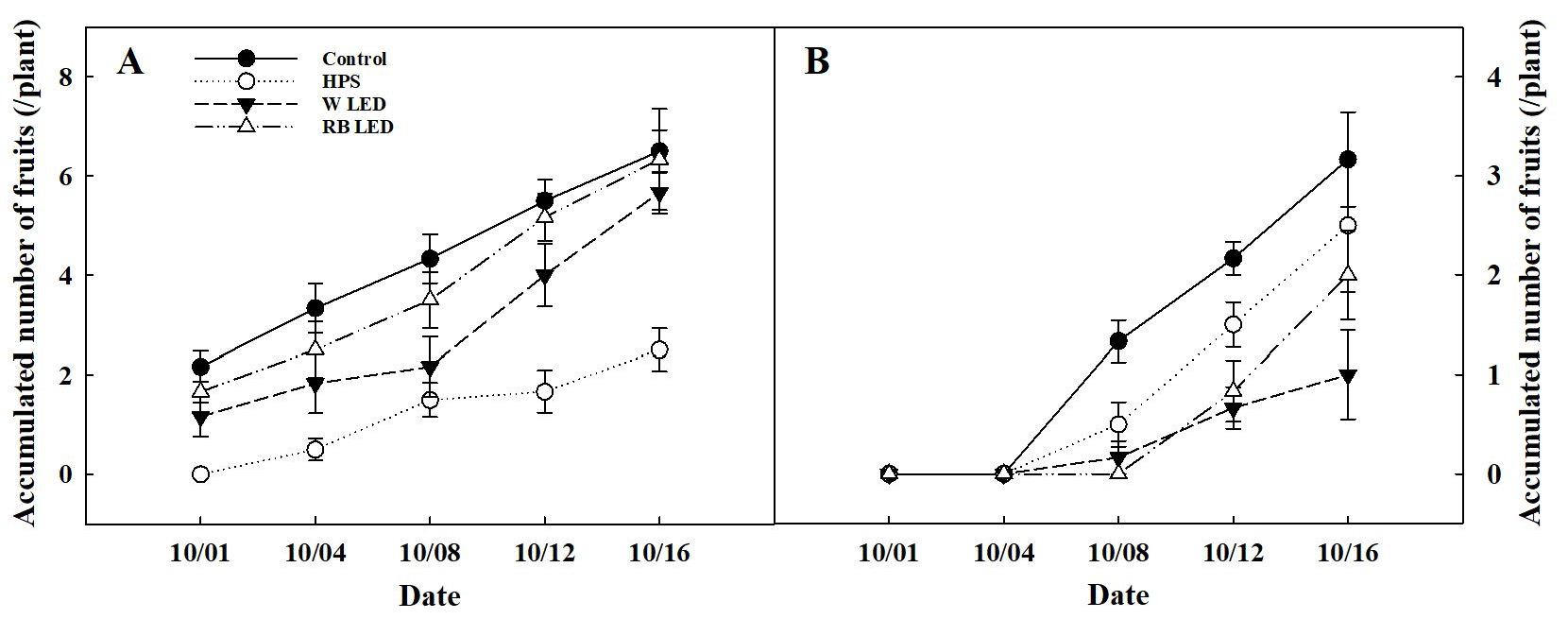
Fig. 6.
Accumulated number of cucumber fruits as affected by shading level under different supplemental lighting sources. A, 0% shading level and B, 50% shading level. Refer to Fig. 4 for details on spectral distribution of supplemental lighting sources.
따라서, 본 연구에서는 하절기 차광률 50% 수준의 차광 처리가 오이의 초장, 엽장, 엽폭, SPAD 값을 증가시켰으나, 보광 처리에 따른 오이의 생육 증진 효과는 RB LED로 보광 하였을 때 경경이 유의하게 증가한 것을 제외하고 유의적인 차이가 없었다. 또한, 과경, 과실 생체중과 건물중은 차광, 보광에 따른 차이가 없었고, 누적 과실 개수는 보광 대조구가 많았다. 위의 결과를 종합하였을 때 하절기 오이 재배 시 50% 수준의 차광 처리는 오이의 생육을 유의하게 향상시켰다. 또한, 보광 대조구에서 생육과 과실 특성이 좋았다. 따라서 하절기 오이 재배에 보광 기술을 적용하기 위해서는 적절한 차광 수준과 보광 시간, 광도 등을 고려한 추가적인 연구가 필요할 것으로 판단된다.
