

서 론
포도의 품질은 맛, 크기, 색 등의 다양한 요인에 의해 판단되며, 그 중에서 과피색은 소비자가 품질을 판단하 는데 있어 중요한 요인으로 여겨지고 있다. 포도의 과피 색은 안토시아닌이라는 색소의 종류 및 농도로 결정되며, 품종 고유의 안토시아닌 조성은 유전적인 요인에 의해 결정된다(Castellarin과 Gaspero, 2007). 하지만, 같은 품 종에 대해서도 온도나 광 등의 환경 조건에 의해 안토 시아닌 조성이 영향을 받을 수 있으며(Tarara 등, 2008), 일반적으로 고온보다는 저온 조건에서 생합성이 촉진된 다(Yamane 등, 2006).
최근 여름철 폭염 및 열대야 발생 빈도의 증가로 인해 포도의 과피색 불량 문제가 나타나고 있으며, ‘거봉’ 포 도의 경우 변색기 고온에 의해 착색이 멈춘 뒤 다시 회 복되지 못하는 문제점이 발생하고 있다(Ryu 등, 2018). 포도의 착색을 증진시키기 위해 환상박피, 토양수분 조 절, 적엽 등의 재배적인 조치에 대한 연구가 수행되어 왔지만(Castellarin 등, 2007;Koshita 등, 2011;Lemut 등, 2011), 과방에 식물 호르몬 abscisic acid(ABA)를 살 포하는 것이 가장 효과적인 것으로 알려졌다(Jeong 등, 2004;Cantin 등, 2007). 실제로 ABA는 포도의 성숙을 조절하는 가장 중요한 식물 호르몬으로(Pilati 등, 2017), 과피에서 변색기를 기준으로 함량이 증가하며(Wheeler 등, 2009), 과방에 ABA를 직접 살포 시 안토시아닌 생 합성 유전자의 발현을 증가시켜 착색을 증진시킨다고 알 려져 있다(Jeong 등, 2004).
하지만 고온 조건에서 과피의 착색이 정지하여 회복되 지 못하는 원인에 대해서는 아직 정확하게 밝혀진 바가 없으며(Ryu 등, 2018), 고온으로 인한 착색 정지에 관여 하는 식물 호르몬 및 유전자 발현 분석을 통한 근본적 인 원인 구명이 필요한 상황이다. 따라서, 본 연구에서 는 변색 초기의 고온에 의한 ‘거봉’ 포도의 착색을 분석 하고, 착색 정지의 원인을 구명하기 위해 시기별로 ABA, gibberellic acid(GA) 농도 및 대사 관련 유전자 발현을 함께 분석하였다.
재료 및 방법
1. 공시 작물 및 온도 처리 방법
전라북도 완주군 국립원예특작과학원내의 유리온실에 서 포트에 재식한 3년생 ‘거봉’ 나무를 대상으로 변색기 고온 처리구와 정상구의 과피색 변화, 식물호르몬, 유전 자발현 정도를 비교하였다. 고온 처리구는 7월 16일(변 색기, 전체 송이의 10%가 착색)부터 7월 25일까지 대조 구에 비해 6°C 높은 온도를 처리하였으며, 대조구는 완 주 지역의 30년 평균 기온을 시간 별로 적용하였다. 시 험구당 6주씩 처리하였고, 한 주당 4-5송이(송이당 성엽 14매 이상 확보)의 포도를 착과시켜 과실 품질 및 물질 분석에 이용하였다. 온도 처리가 종료된 이후에는 대조 구와 고온구의 온도를 모두 완주지역 7, 8월의 30년 평 균 기온으로 적용하여 착색의 진행을 관찰하였다.
2. 과실 품질 조사
과실의 품질은 10월 4일에 일괄 수확하여 과방중, 과 립수를 조사하였고, 송이 당 10개의 과립을 선정하여 양 쪽 적도면을 대상으로 색차계(CR-300, Minolta, Osaka, Japan)를 이용하여 색차값을 측정하였다. 동일한 시료를 대상으로, 과즙을 착즙하여 굴절 당도계(PR-101, Atago, Tokyo, Japan)로 당도를 측정하고 적정 산도(0.1N NaOH, pH 8.1)를 조사한 후 주석산으로 환산하여 표기 하였다. 시기별 과피색의 변화는 농촌진흥청에서 제시한 ‘거봉’의 성숙기 판정용 칼라차트(Park 등, 2004)를 기준 으로 조사하였다.
3. 과피의 안토시아닌 정량 분석
과피의 안토시아닌 함량을 분석하기 위해 수확기 송이 에서 시료를 채취하였다. 안토시아닌의 추출 및 분석은 Ryu 등(2018)의 방법을 활용하였고, HPLC(1100 Series, Hewlett-Packard, Germany)로 분리 및 정량 분석하였다. 안토시아닌 분리에 이용한 컬럼은 YMC-Pack Pro C18 RS(250 × 4.6mm, 5mm, YMC, Japan)이며, 40°C에서 41 분 동안 분석하였다. 안토시아닌의 함량은 518nm의 파 장에서 malvidin 3-glucoside(Extrasynthese, France)의 표 준품을 이용해 작성한 표준곡선으로 정량하여 mg·g-1의 단위로 나타내었다.
4. 과피의 식물호르몬 정량분석
식물 호르몬 중 ABA의 추출 및 정량분석은 Ryu 등 (2016)의 실험 방법을 활용하였다. GA1은 50% MeOH 을 추출 용매로 활용하였고, 내부표준물질인 d2-GA1을 4ng씩 첨가시켜 4°C 암조건에서 15시간 동안 추출하였 다. 추출된 용매는 MeOH로 전처리된 sep-pak C18 카 트리지에 통과시켜 40°C의 온도로 감압 농축시킨 뒤 MeOH에 재용해시켜 분석하였다. ABA 및 GA1의 함량 은 ng·g-1의 값으로 나타내었으며, 3반복으로 분석하였다. 식물 호르몬의 분석을 위한 HPLC 기기 조건은 Ryu 등 (2016)의 실험 방법과 동일하였고, 정량분석은 ion trap MS detector(Finnigan LXQ, Thermoelectron, Marietta, USA)가 부착된 HPLC(Nanospace SI-2, Shiseido, Japan)를 이용하였다. 내부표준물질과 endogenous peak 면적의 비 율을 기본으로 정량이 이루어졌으며, ABA는 263→153m/z, d6-ABA는 269→159m/z, GA1은 347.1→303.1m/z, d2-GA1 은 349.2→305.2m/z를 multiple reaction monitoring (MRM) 조건으로 각각 설정하여 정량하였다.
5. RNA 추출 및 qRT-PCR을 통한 유전자 발현 분석
과피의 ABA, GA 대사 관련 및 안토시아닌 생합성 관 련 유전자의 발현은 RNA를 추출하여 quantitative realtime polymerase chain reaction(qRT-PCR)을 이용하여 분석 하였으며, RNA 추출 및 cDNA 합성 조건은 Ryu 등(2017) 의 실험 방법을 이용하였다. qRT-PCR은 LightCycler® 480 Real-time PCR System(Roche Diagnostics, Mannheim, Germany)으로 수행하였고, reaction mixture는 TB GreenTM premix Ex TaqTM (Takara, Tokyo, Japan)를 이용하여 제작 하였다. 최종 reaction volume은 제조사의 권장 사용법에 따라 20mL로 설정하였다. 실험에 이용한 primer의 염기 서열은 Table 1과 같다.

결 과
1. 과피의 착색 및 과실품질
변색기(7월 16일, 만개 후 52일)부터 10일 동안의 고 온에 의해 ‘거봉’ 포도의 과피색 불량이 나타났다(Fig. 1). 성숙기 판정용 칼라차트를 기준으로 과피색을 조사 한 결과, 고온처리 직후 대조구에 비해 고온구의 과피 착색이 진행되지 못하였고, 이러한 경향은 ‘거봉’의 상업 적 수확기인 만개 후 약 100일(변색 48일 후)까지 유지 되었다. ‘거봉’의 일반적인 과피색인 흑청색을 나타내는 대조구와 달리, 고온구의 송이에는 녹색 및 밝은 적색을 띠는 과립이 섞여있었다.
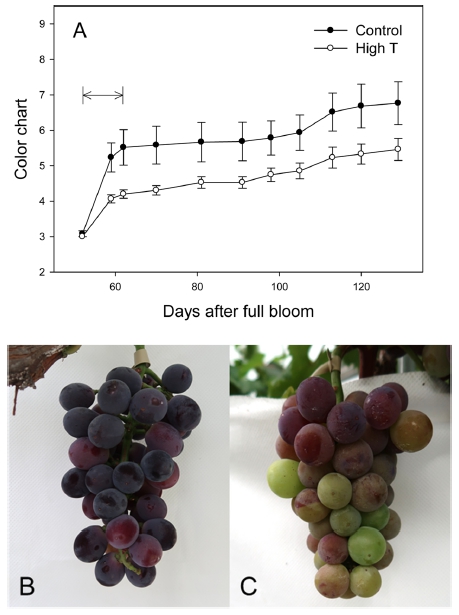
Fig. 1
Coloration of ‘Kyoho’ grape berry skins under control and high temperature (High T) conditions. Arrow means the High T treatment period. (A) Changes in color chart of ‘Kyoho’ grape berry skins. Vertical bars indicate the standard errors (n = 4). (B, C) Grape bunches under control (B) and High T (C) conditions, respectively. Photographs were taken at 101 days after full bloom.
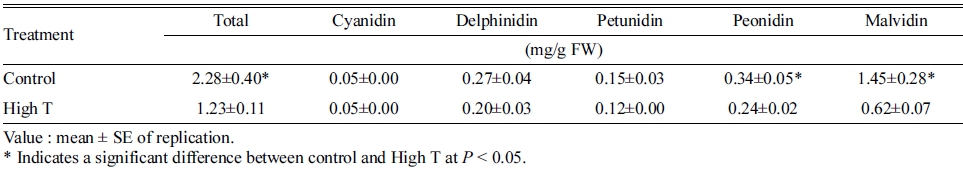
실제 과피의 안토시아닌 함량을 비교한 결과, 고온 처 리에 의해 총 안토시아닌 함량이 대조구의 54% 수준으 로 감소하였다(Table 2). 안토시아니딘의 그룹별로 비교 하면 malvidin(Mal), peonidin(Peo)이 대조구에 비해 유 의하게 감소하였으며, delphinidin(Del), petunidin(Pet), cyanidin(Cya)은 고온 처리에 의해 감소하지 않았다.
Table 2. Anthocyanin contents in berry skins of ‘Kyoho’ grape under control and high temperature (High T) conditions at 132 days after full bloom.
 |
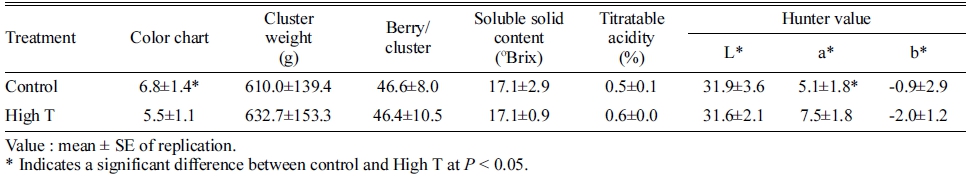
고온 처리에 의한 과실품질을 비교한 결과, 착색과 관 련된 항목을 제외한 나머지 과실 품질은 고온에 의해 영향을 받지 않았다(Table 3). 칼라차트로 비교한 과피의 착색도는 고온에 의해 감소하였고, Hunter 값 중 붉은색 의 정도를 나타내는 a*값이 유의하게 증가하였다. 과중, 과실의 당도 및 산도는 고온 처리에 의해 영향을 받지 않았다.
2. 식물호르몬 ABA 및 GA 함량
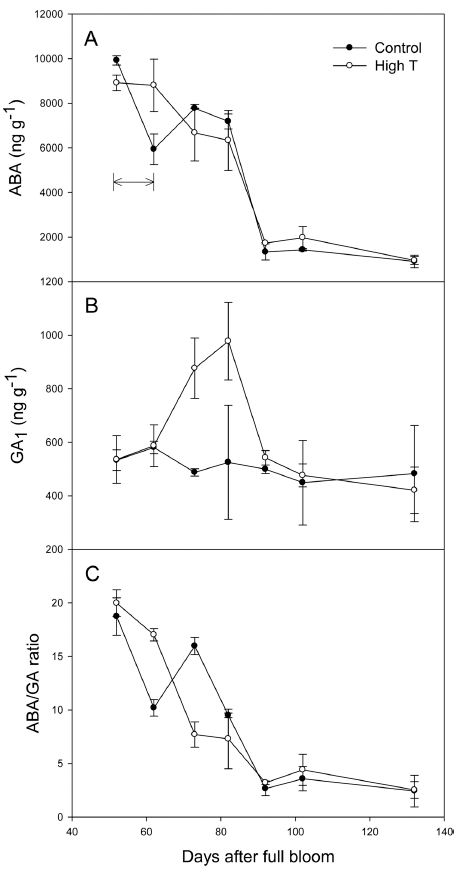
고온에 의해 착색이 정상적으로 진행되지 못하고 정지 된 원인을 구명하기 위해 과피의 식물 호르몬 ABA와 GA의 함량을 분석하였다(Fig. 2). 과피의 ABA 함량은 변색기(만개 후 52일) 근처에 높게 유지되었다가 만개 후 92일을 기준으로 급격히 감소하는 경향을 보였으며, 고온에 의해 감소하지 않았다. 처리 직후에는 오히려 대 조구에 비해서 높은 경향을 보였으며, 처리 종료 10일 후 대조구와 비슷한 수준으로 감소하였다. GA는 전체 활성 형 GA 중에서 포도 과실에 가장 많은 양으로 존재하는 것으로 확인된 GA1을 분석하였다(Ryu, unpublished data). 고온처리 종료 10일 후부터 과피의 GA함량이 대조구의 약 2배로 증가하였으며, 30일 후 다시 대조구의 수준으 로 감소하였다. ABA와 GA의 비율을 분석한 결과, 고온 처리 직후 ABA/GA 값이 일시적으로 높아졌다가 만개 후 73일에 대조구에 비해 감소하였다(Fig. 2).
3. 안토시아닌 생합성 유전자 발현
고온이 안토시아닌 합성의 어느 단계에서 영향을 미치 는지 분석하기 위해 시기별 안토시아닌 생합성 유전자의 발현을 분석하였다(Fig. 3). 안토시아닌 생합성을 조절하 는 초기 생합성 유전자(Early biosynthetic genes, EBG) 중에서 과피의 착색과 동일한 경향을 보이며 발현이 조 절된다고 알려진 CHS3, F3H1과 후기 생합성 유전자 (Late biosynthetic genes, LBG) 중에서 DFR, UFGT의 발현을 분석하였다(Azuma 등, 2012). CHS3는 고온에 의해 처리 종료 20일 후까지 대조구에 비해 발현이 높 았으며, 이후 대조구와 비슷한 수준으로 감소하였다. F3H1와 DFR은 고온에 의해 발현이 영향을 받지 않았 다. 안토시아닌 생합성의 가장 마지막 단계를 조절하는 UFGT는 고온 처리 직후 대조구에 비해 발현이 감소하 였으며, 처리 종료 20일 후에는 다시 대조구의 수준으로 증가하여 수확기까지 차이를 나타내지 않았다.
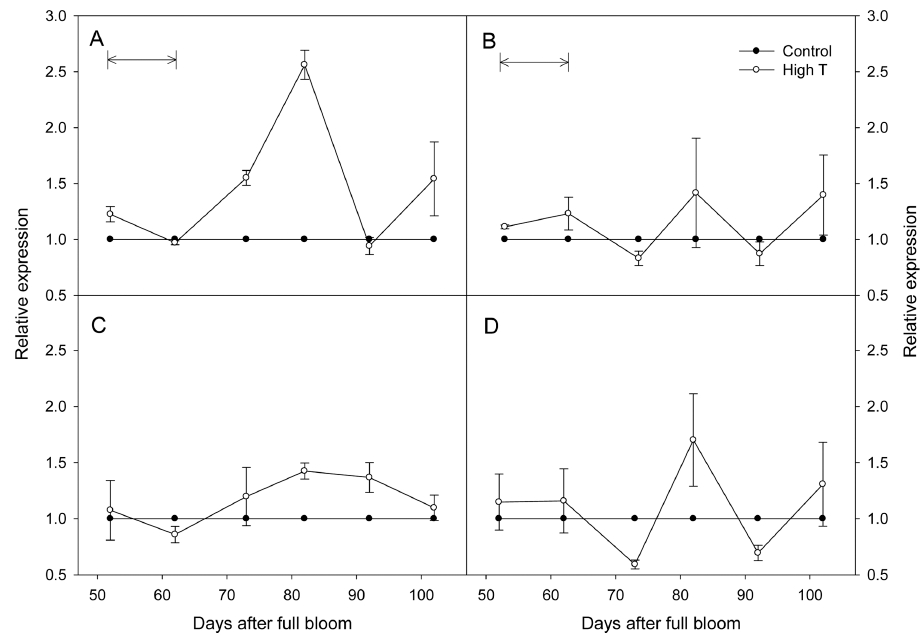
Fig. 3
Fold changes in the expressions of anthocyanin biosynthetic genes, CHS3 (A), F3H1 (B), DFR (C), and UFGT (D) in ‘Kyoho’ grape berry skins under control and high temperature (High T) conditions. Arrows mean the High T treatment period. The expressions were normalized against the expression of the grape actin gene. Vertical bars are the standard errors of the means (n = 4).
4. ABA 및 GA 대사 관련 유전자 발현
고온에 의해 과피의 호르몬 농도가 변화했던 원인을 분석하기 위해, 시기별 ABA 및 GA의 대사와 관련된 유전자의 발현을 분석하였다. ABA의 생합성, 분해를 조 절하는 NCED1, ABA8’OH2의 발현을 분석한 결과, 고 온에 의해 두 유전자의 발현이 모두 대조구에 비해 감 소하지 않았다(Fig. 4). GA에 대해서는 생합성을 유도하 는 GA20ox1/2와 불활성화를 조절하는 GA2ox1/2의 시기 별 발현을 분석하였다(Fig. 5). 실험 결과, 고온 처리와 동시에 GA20ox1의 발현이 대조구에 비해 증가하였고, 처리 종료 10일 후까지 높게 유지되었다. GA20ox2는 고온에 의해 영향을 받지 않았으며, 특별한 경향을 보이 지 않았다. GA2ox1/2는 고온 처리 직후 모두 발현이 감 소하였으며, 처리 종료 10일 후까지 대조구에 비해 낮게 유지되었다.
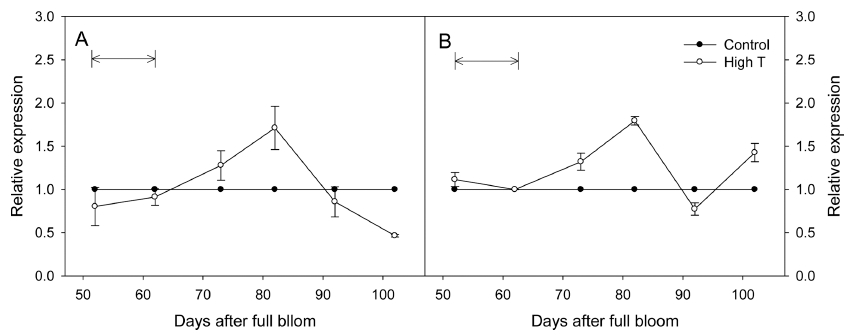
Fig. 4
Fold changes in the expressions of ABA metabolic genes, NCED1 (A), and ABA8’OH2 (B) in ‘Kyoho’ grape berry skins under control and high temperature (High T) conditions. Arrows mean the High T treatment period. The expressions were normalized against the expression of the grape actin gene. Vertical bars are the standard errors of the means (n = 4).
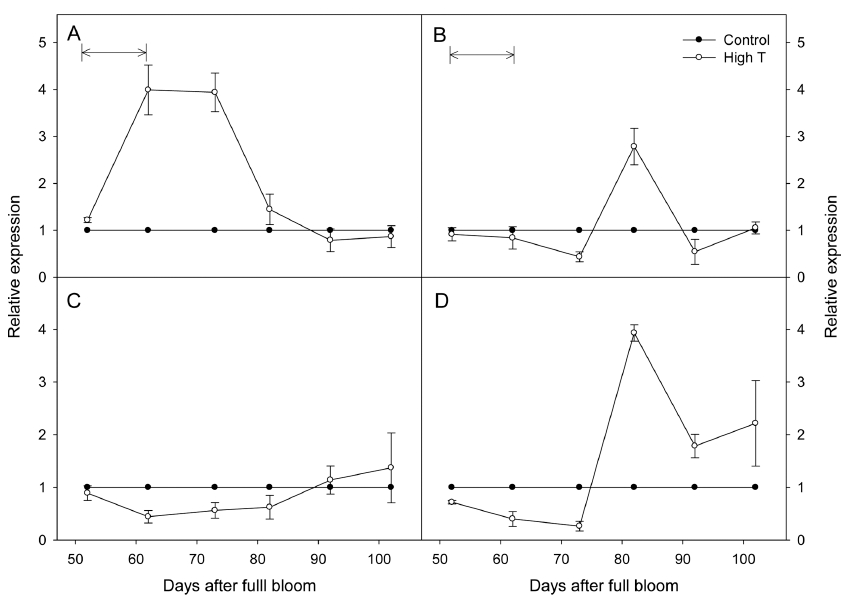
Fig. 5
Fold changes in the expressions of GA metabolic genes, GA20ox1 (A), GA20ox2 (B), GA2ox1 (C), and GA2ox2 (D) in ‘Kyoho’ grape berry skins under control and high temperature (High T) conditions. Arrows mean the High T treatment period. The expressions were normalized against the expression of the grape actin gene. Vertical bars are the standard errors of the means (n = 4).
고 찰
1. 변색기 고온에 의한 과피색 불량
변색기부터 10일 동안의 고온에 의해 ‘거봉’ 포도의 과피색이 불량하게 나타났으며(Fig. 1), 그 원인은 과피 의 총 안토시아닌 함량이 감소했기 때문이었다(Table. 2). 과실의 당도는 일시적인 고온에 의해서 영향을 받지 않았으며, 품질의 변화 없이 과피의 착색만 정상적으로 진행되지 못하고 멈춘 현상을 나타내었다(Table. 3). 과 피의 색차값 중 hunter a*값이 고온에 의해 증가한 것은 착색이 정상적으로 진행되지 못했다는 것을 의미한다. ‘거 봉’과 같은 흑청색 품종의 경우, 변색 초기에는 밝은 붉 은색을 나타내지만 성숙이 진행될수록 안토시아닌의 함 량이 높아지며 자흑색을 띠게 된다. Park 등(2010)은 변 색기를 기준으로 ‘거봉’ 과피의 착색도를 분석했을 때, 붉은 색을 나타내는 hunter a*값이 변색 2주 후까지 증 가하다가 다시 감소한다고 하였으며, 최종적으로 안토시 아닌 함량이 낮은 송이일수록 높은 hunter a*값을 나타 낸다고 하였다. 본 연구의 결과에서도 고온에 의해 과피 의 안토시아닌 함량이 감소하였고, 결과적으로 대조구의 과립보다는 밝은 적색을 띠며 hunter a*값이 높은 결과 를 나타내었다.
‘거봉’의 성숙기 고온에 의한 과피 착색에 대해서는 많은 연구가 이루어져 왔으며, 본 연구의 결과는 변색기 부터 수확기까지 고온에 의해 과피의 안토시아닌이 감소 하였다는 Mori 등(2004)과 Shinomiya 등(2015)의 결과 와 일치하였다. 하지만, 고온에 의해 과실로의 광합성 동화산물의 이동이 감소하였다는 Mori 등(2004)의 결과 및 과실의 당 함량이 유의하게 증가하였다는 Pastore 등 (2017)의 결과와 달리, 본 연구에서는 과실의 당도가 고 온에 의해 영향을 받지 않았다(Table 3). 이는 고온에 노 출시켜 실험을 진행했던 기간이 달랐기 때문으로 판단되 며, 본 연구에서는 변색 초기에 10일이라는 비교적 짧은 기간 동안 처리가 진행되었기 때문에 고온이 최종적으로 과실의 당함량에 영향을 미치지 않은 것으로 판단되었다.
2. 안토시아닌 조성 및 생합성 변화
과피의 착색이 정상적으로 진행되지 않았던 원인을 분 석하기 위해 안토시아니딘의 그룹별로 분석한 결과, 전 체 5가지 그룹 중에서 Mal과 Peo만이 고온에 의해 영향 을 받아 감소하였다(Table. 2). 이 두 그룹은 안토시아니 딘의 종류를 결정하는 특징적인 구조(flavilium)에서 B ring에 methoxyl group을 가지며, 외부 스트레스에 의한 산화 반응에 비교적 안정적이라고 알려져 있다(He 등, 2010). 또한, 대조구의 과피에서 총 안토시아닌 함량에 대해 각 안토시아니딘 그룹의 함량을 비교했을 때 Mal 과 Peo가 전체의 약 80%를 차지하고 있었으며(Table. 2), 이는 기존의 Li 등(2013) 및 Ryu 등(2018)의 보고와 도 일치하는 결과였다. 따라서 변색 초기의 고온에 의한 ‘거봉’의 과피색 불량은 기존에 합성된 안토시아닌이 분 해되었다기 보다는, 안토시아닌의 생합성이 전체적으로 억제되었기 때문이라고 판단되었다. 실제 과피 내 안토 시아닌 생합성 유전자 중 UFGT의 발현이 고온에 의해 억제되었으며(Fig. 3), 이는 생육기 고온 조건에서 과피 의 UFGT의 발현이 감소하여 착색이 억제되었다는 De Rosas 등(2017)의 보고와도 일치하는 결과였다.
3. 식물호르몬 농도 변화와 과피색 불량
포도의 착색은 다양한 식물 호르몬의 변화에 의해 조 절되는데, 그 중에서 ABA에 대해 가장 많은 연구가 이 루어지고 중요한 역할을 한다고 알려져 있다(Fortes 등, 2015). 고온 조건에서 포도 과피의 착색 및 ABA 함량 에 대해서는 다양한 의견이 제시되고 있는데, Koshita 등(2007)과 Azuma 등(2012)은 고온에 의해 과피의 ABA 함량이 감소하였으며 그로 인해 안토시아닌 함량 이 감소했다고 보고하였다. 반대로 Shinomiya 등(2015) 과 Carbonell-Bejerano 등(2013)은 고온에 의해 ABA의 함량이 오히려 증가하였지만, 그럼에도 불구하고 안토시 아닌 함량이 감소했다고 보고하였다.
본 실험에서도 변색 초기의 고온에 의해 과피의 ABA 함량이 영향을 받아 감소하지 않았으며(Fig. 2), 생합성 을 조절하는 NCED1 및 ABA8’OH2 유전자의 발현 역 시 고온에 의해 감소하지 않았다(Fig. 4). 기존 문헌과 마찬가지로 대조구와 비슷한 수준의 ABA 함량이 안토 시아닌의 축적으로 이어지지 못했던 이유는, ABA가 다 른 호르몬과의 비율, 특히 GA와의 균형을 통해 안토시 아닌 생합성을 조절하기 때문으로 추측되었다. 애기장대 의 경우, 안토시아닌 생합성은 다양한 호르몬의 상호작 용에 의해 결정된다고 밝혀져 있으며(Das 등, 2012), 당 에 의한 안토시아닌 축적 경로의 경우 ABA가 안토시아 닌의 생합성을 촉진시키는 역할을 하는데 비해 GA는 그것을 억제하는 역할을 한다(Weiss와 Ori, 2007;Loreti 등, 2008). 실험의 결과, 변색 초기의 고온에 의해 GA 생합성 유전자 중 GA20ox1의 발현 증가 및 불활성화 관련 유전자 GA2ox1/2의 발현이 감소하였고(Fig. 5), 과 피의 GA 함량이 증가하였다(Fig. 2). 이로 인해 ABA/ GA 비율이 고온에 의해 일시적으로 감소하였으며(만개 후 73일), 이 경향이 실제 과피의 안토시아닌 생합성을 조절하는 UFGT의 발현 패턴과 유사하였다(Fig.2, 3). 따 라서, 포도 과피의 안토시아닌 생합성 역시 ABA의 절 대적인 함량보다는 ABA와 GA의 비율에 의해서 조절될 가능성을 확인하였다.
변색 초기의 고온으로 인해 ‘거봉’ 포도의 착색이 정 지된 원인은 과피의 안토시아닌 생합성이 고온에 의해 억제되었기 때문이었고, 안토시아닌 생합성은 ABA의 함량보다는 고온에 의한 GA의 증가와 그에 따른 ABA/ GA값의 감소에 의해서 조절되는 것으로 추정되었다. 여 름철 이상 고온으로 인한 과피의 착색 불량을 경감시키 기 위해서는 안토시아닌 생합성을 촉진시키는데 있어 다 양한 식물 호르몬의 관계를 명확히 구명할 필요가 있으 며, 본 연구에서 다루었던 활성형 ABA나 GA의 정량분 석뿐만 아니라 고온에 의한 실시간 호르몬 대사 변화에 대한 연구가 추가로 필요하다고 판단된다.
