

서 론
수박(Citrullus lanatus Thunb.)은 전 세계적으로 널리 재배되는 대표적인 여름 과채류로 국내에서도 고온기 고품질 과실 생산을 위한 시설 재배 비중이 2023년 기준 9,330ha로 점차 확대되어 왔다(KOSIS, 2023). 특히 1인 가족, 소비자의 기호 변화 등에 따라 소과종 씨 없는 수박의 수요가 증가하고 있으며(KREI, 1995), 이와 함께 안정적인 착과 및 수량 확보를 위한 재배기술 개발의 중요성이 강조되고 있다. 하지만 씨 없는 수박은 공동과와 같은 비상품과 발생이 많고 중심부와 주변부의 당도 차이가 크다는 단점이 있는 것으로 알려졌다(Diezma-Iglesias 등, 2004; Phat 등, 2015). 그리고 발아율이 낮아 종자 가격이 비싸고 기상환경이 좋지 않을 때 착과율이 저하되기 때문에 생식 생장 초기의 환경 조건 관리가 매우 중요하다.
시설 환경에서 작물의 생육과 생식 생장은 주로 온도, 광, 습도 등의 환경적 요인에 의해 크게 영향을 받으며, 특히 광 환경은 식물의 광합성 작용을 통한 생장량 증가 및 과실 품질에 영향을 미치는 요인으로 작용한다(Heuvelink 등, 2004; Jeong 등, 2009a, 2009b; Marcelis 등, 2004). 암꽃 착생은 식물체의 에너지 축적 및 광 환경 변화에 민감하게 반응하며 착과 수와 수확량에 직접적인 영향을 미치는 중요한 요인 중 하나이다. 온실 환경에서는 계절적 요인이나 기상 조건에 따라 광량이 변화하는데, 특히 겨울철 자연광이 부족할 경우 수박 모종의 생육 품질이 저하되고 재배 기간이 길어질 수 있다(Wei 등, 2020). 이와 같이 광 환경은 작물의 생장, 발달 및 수량 등에 중요한 요인으로 지속되는 흐린 날씨는 저광 스트레스를 유발할 수 있다(Ma 등, 2025). 이를 보완하기 위해 차광망이나 보광등을 활용한 광 조절 기술이 도입되고 있다. 재배과정 중 보광 처리는 광합성율, 생장량, 암꽃 발생량 등을 유의하게 증가시킬 수 있다(Chamchum 등, 2023; Hossain 등, 2025). 하지만 실제 국내 수박 재배 현장에서 차광 및 보광 처리의 효과를 정량적으로 분석한 연구는 매우 제한적이다.
따라서 본 연구는 차광과 보광을 통한 광량 수준의 차이가 수박의 생장, 암꽃 발생 및 과실 특성에 미치는 영향을 알아보고자 수행되었으며, 향후 씨 없는 수박의 안정적인 생식 생장과 수량 증대를 위한 광 환경 기반 생육 모델 수립 및 보광 수준 설계에 기초자료를 제공할 것으로 기대된다.
재료 및 방법
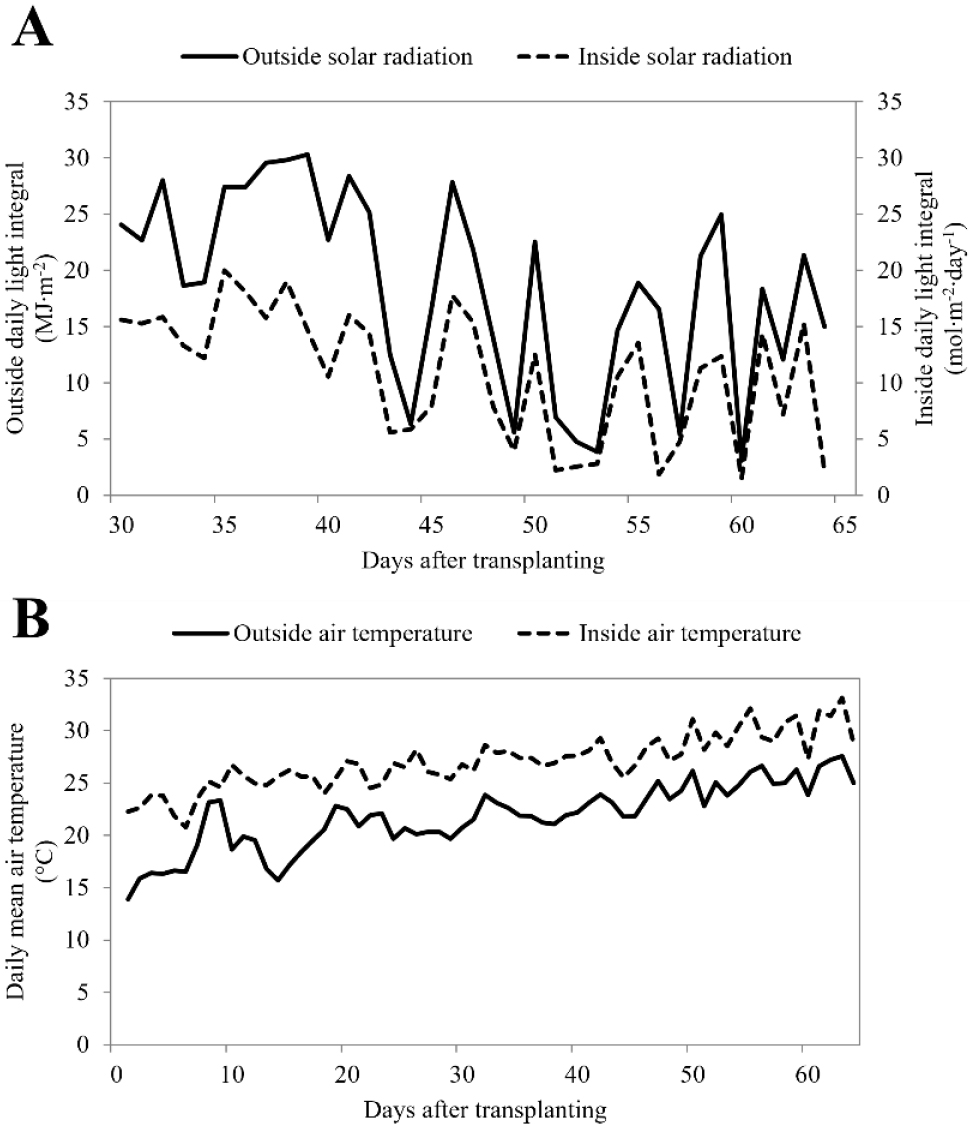
본 실험은 전라북도농업기술원 수박시험장 내 위치한 무가온 연동형 온실(폭 8m, 측고 4.5m, 동고 5.7m, 피복재 Polyethylene film)에서 수행되었다. 온실은 남북(SN)방향이었고, 행거형 지주를 활용한 줄기 유인 방식(∩형)이었다. 실험 품종은 소과종 수박 ‘블랙보이(Black Boy, triploid, Partner Seed Inc. Korea)’로 2023년 5월 9일에 정식되었다. 이후 식물체 당 3줄기를 키우고 이 중 2줄기에 과실 1개씩 착과를 목표로 수분 및 관리하였다. 생육 기간 동안 사용할 양액(100배액 1,000L)을 조제하기 위해 사용된 비료별 사용량은 A액에는 KNO3 25.25kg, (CaNO3)2·4H2O 70.8kg, Fe-EDTA 2kg이었고, B액에는 KNO3 25.25, NH4H2PO4 11.5kg, MgSO47H2O 36.9kg, H3BO3 300g, MnSO4·4H2O 200g, ZnSO4·7H2O 22g, CuSO4·5H2O 5g, Na2MoO4·2H2O 2g이었다. 양액 공급 시 EC는 활착기 1.2, 영양생장기 1.5, 착과 이후 비대기 1.8, 수확기 2.05dS·m-1, pH는 6.5로 조정하여 드리퍼를 이용하여 점적 관수하였다. 광량 수준은 자연광(Natural Light, NL), NL의 차광 70 및 30%, NL+보광(Supplemental Light, SL) 120 및 240PPFD로 5처리구였다. 보광등은 LED White : Red = 3 : 1로 조합된 것으로 식물체 상부 기준 약 150cm 위 온실 천장 구조물에 설치되었다. 보광 처리구의 광량은 식물체에 도달 수치이며, NL이 5분동안 평균 400PPFD 이하 또는 이상 시 점등 또는 소등되도록 설정하였다(Fig. 1).

온실 내부 환경 요소 수집 기간은 정식한 날부터 약 64일 동안이었으며, 기온, 광도(PPFD)는 각 요소의 센서가 장착된 환경계측기(aM-31, Wisesensing, Korea)를 30분 간격으로 측정 및 자동 저장하도록 설정하여 대조구 재식 라인 옆 지주 높이(온실바닥으로부터 약 1m)에 설치하였다. 그리고 온실 외부의 환경 요소는 기상청(Korea Meteorological Administration, KMA)에서 제공한 데이터를 기반으로 작성하였다. 적산온도는 30분 간격으로 수집된 기온을 일평균기온으로 계산한 후 누적하였고, 누적광량은 30분 간격으로 측정된 NL처리구의 광도(PPFD) 값에 차광처리구들은 차광률을 곱하여 빼주고 보광처리구들은 더하여 계산하였다. 그리고 이를 다음 Eq. (1)을 이용하여 일누적광량(Daily Light Integral, DLI)으로 계산한 후 시간의 경과에 따라 누적하여 활용하였다.
DLI : 일누적광량(mol·m-2·day-1)
Pi : PAR의 순간 강도(μmol·m-2·s-1)
Tr : Pi의 1시간 동안 누적광량(Pi×1,800초×2)
DH : 일조시간 (hrs·day-1)
식물체 생육조사는 각 처리구당 7주를 반복수로 하였고, 환경 요소 수집과 같은 기간동안 일주일 간격으로 진행하였다. 생육조사의 항목은 과실 수를 포함한 암꽃 착생 수, 초장, 마디 수, 엽장, 엽폭 그리고 식물체에 착과된 과실의 과장 및 과폭이었으며, 과실의 체적은 과장과 과폭을 측정한 결과 과형지수가 1.0에 근접한 과형으로 판단되어 구의 체적공식(V=4/3πr3)을 이용하여 계산하였다. 이때 반지름(r)은 과장과 과폭의 평균값의 절반으로 하였고 원주율(π)은 3.14159로 하였다. 그리고 파괴조사는 엽면적, 생체중, 건물중, 과실 무게, 당도를 측정 후 평균하였다.
조사된 데이터는 SPSS 프로그램(Version 29.0 SPSS Inc., Chicago, USA)을 통해 분산분석(One-way ANOVA)되었으며, 각 그래프는 Excel 2021(office 365, microsoft Crop., Canada)을 이용하여 작성하였다.
결과 및 고찰
정식한 날부터 64일까지의 실험 기간 동안 온실 외부 및 내부의 일평균기온 및 일누적광량의 변화를 확인하기 위해 시계열로 나타내었다(Fig. 2). 실험 기간 동안 외부 및 내부의 일누적광량은 각각 평균 18.5 ± 8.4MJ·m-2·day-1, 10.9 ± 5.7mol· m-2·day-1였다. 그리고 외부 및 내부의 일평균기온은 각각 평균 23.8 ± 1.9, 28.7 ± 1.9℃였다. 특히, 일누적광량의 일별 편차가 매우 컸는데 이는 운량, 강수 등 외부 요인에 따른 것이며(KMA, 2023), 온실 내부는 외부보다 평균 기온이 약 최소 1.3°C, 최고 10.0°C 높게 유지되었고, 두 요소 모두 외부의 변화를 따르는 경향이었다.
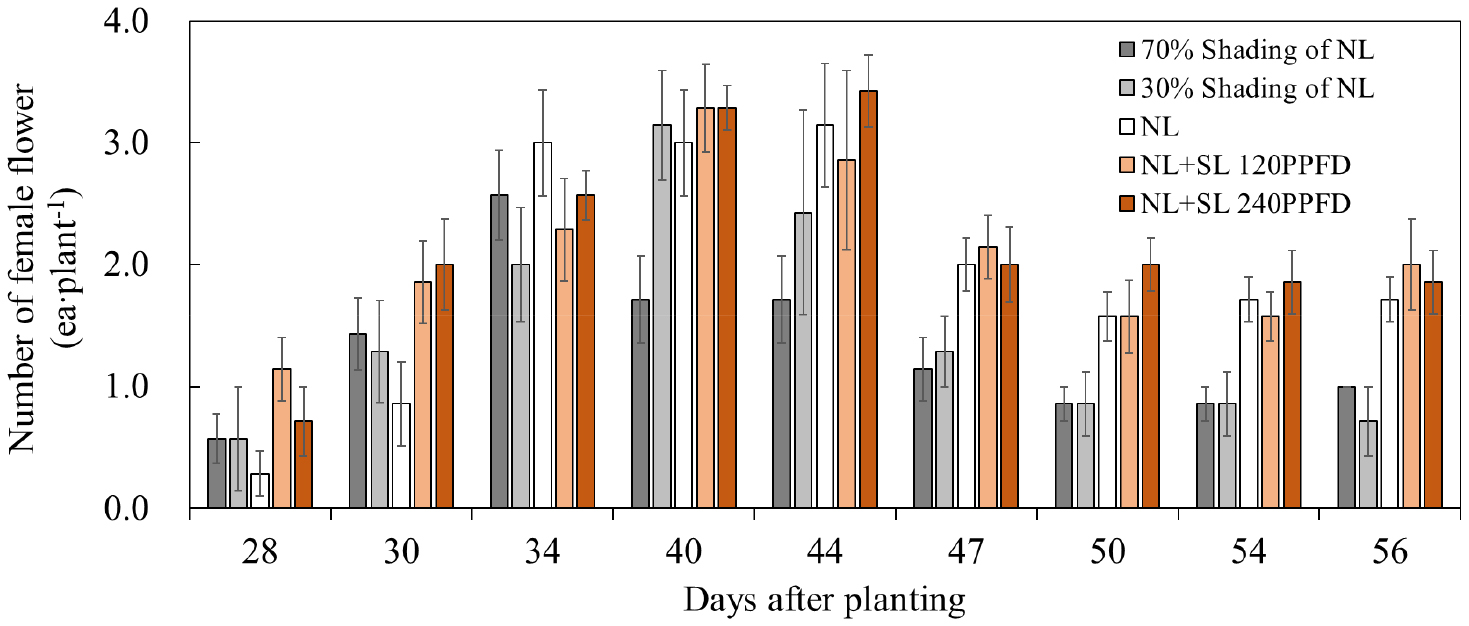
실험 기간 동안 각 처리구별 과실을 포함한 암꽃 발생 수를 조사한 결과(Fig. 3), 70% 차광 처리는 전체 기간 동안 주당 암꽃 발생 수가 가장 적었으며, 30% 차광은 70% 차광보다 높았으나 NL 및 보광 처리보다 낮은 값을 보였다. 그리고 대조구는 전체 기간 동안 안정적인 암꽃 발현을 나타내었다. 또한 보광 처리구에서 공통적으로 정식 후 약 34-44일 구간에서 증가량이 가장 많았고, 점차 줄어드는 경향이었다. 특히 보광 처리구 중 NL + SL 240PPFD에서의 암꽃 발현 수가 가장 높았는데 정식 후 40일 전후에는 식물체당 평균 3.5개 이상의 암꽃 발생이 확인되었다. 이러한 광 환경 수준에 따른 암꽃 발생 수는 광이 과도하게 제한되지 않았을 때 암꽃이 활발하게 발생하는 등 광 환경에 의존하는 경향을 보였지만 통계적으로 유의한 차이를 나타내진 않았다. 특히 보광 처리는 암꽃 착생 수를 증가시키는 데 효과적이었으며, 보광 처리보다 저온 및 단일조건인 차광 처리에서 암꽃 착생을 빠르게 유도하되 광량 부족 시 암꽃이 작아지고 낙화량이 많아진다고 하였는데(ARES, 2025) 본 연구에서도 같은 결과였다. 이러한 결과는 저광 조건에서 암꽃 발생 수가 감소하며(Ma 등, 2025), 이에 따른 착과율 저하가 나타난다는 보고(Marcelis 등, 2004)와 LED 보광 처리가 암꽃 발현 수를 유의적으로 증가시킨다는 결과(Choi 등, 2023; Hossain 등, 2025)와도 일치하는 결과였다.
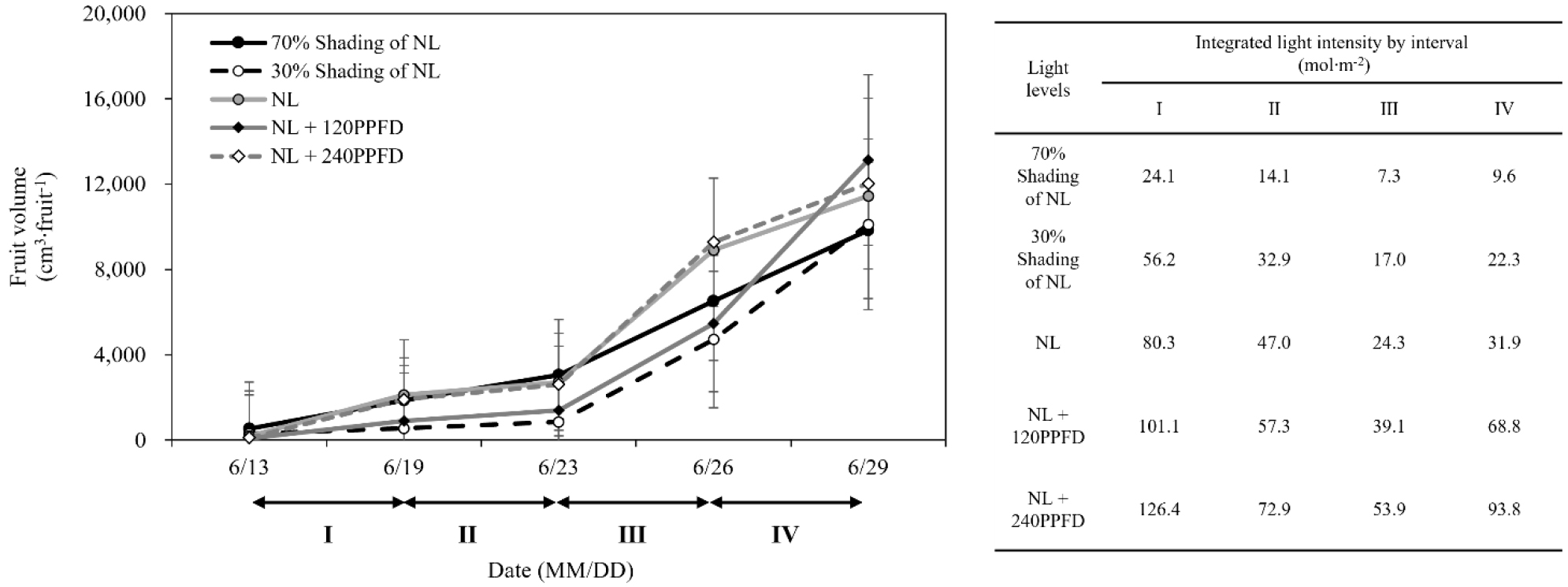
수박의 과실 비대는 일반적으로 이중시그모이드 곡선을 이룬다. 하지만 본 품종은 중소형과로 단일 시그모이드 곡선을 나타내었다(자료 미제시). 이를 기반으로 광 환경 수준에 따라서 착과 후 과실비대 초기의 체적증가와 누적광량에 대하여 알아보았다(Fig. 4). 전체적으로 과실의 최대 비대속도가 변화하는 시점은 모든 처리에서 유사하게 나타내어 광량보다는 온도, 그리고 내부 요인에 의한 발달 단계의 영향이 큰 것으로 판단되었다. I-Ⅱ 동안에는 누적광량의 차이에 따라 과실 체적 증가량이 뚜렷한 관계를 나타내지 않았다. 이후 Ⅲ-Ⅳ 동안 누적광량은 차광 처리구들보다 대조구와 보광처리구들에서 과실비대 속도가 빠른 경향을 나타내었다. 특히, 대조구와 NL + SL 240PPFD는 유사하였고, NL + SL 120PPFD 처리구에서는 그 속도가 가장 빨랐다. 이러한 결과는 포도와 딸기에서 누적광량이 과실의 비대에 영향을 주고(Park 등, 2018) 과실 체적 증가에 광합성 생리 활성 차이가 영향을 준다(Woo 등, 2022)는 기존 연구 결과와 유사한 경향을 나타내었다. 하지만 Ⅲ-Ⅳ 동안 NL 56.2, NL + SL 120PPFD 107.9, NL + SL 240PPFD 147.7mol·m-2과 이들의 과실 체적량 변화를 고려할 때 수박의 과실 비대에는 매우 낮은 광량 조건이 아니라면 이후 증가되는 광량의 영향이 크지 않고 온도의 영향이 더 클 것으로 판단된다(Fukuoka 등, 2009). 적산온도는 과실 발달 속도 주요 요인으로 작용한다(Marcelis 등, 2004).
광 환경 처리에 따른 수박 생육 특성을 분석한 결과(Table 1), 초장은 NL + SL 240PPFD 처리구에서 254.8cm로 가장 길었으며, NL + SL 120PPFD 처리구와 유의적인 차이는 없었다. 반면, 30% 차광 처리구에서는 196.9cm로 가장 짧게 나타나 광 부족이 수박의 줄기 신장을 억제하는 영향을 미친 것으로 확인되었다. 마디 수는 NL + SL 240PPFD 처리구에서 29.2개로 가장 많았으며, 30% 차광 처리구는 21.5개로 가장 적었다. 참외의 경우 같은 온실 내 생육 및 과실 특성이 일사량이 더 많았던 위치에서 증가하는 경향이었는데(Shin 등, 2005), 본 연구에서의 70% 차광 처리구를 제외하고 보광 처리에 의해 초장과 마디 수 증가가 촉진되어 유의한 차이를 나타낸 것은 보광에 의한 기온이 높아진 것(Kwon 등, 2018; Yoon 등, 2023)이 영향을 미쳤을 것이라고 판단된다. 그리고 엽장은 NL + SL 120PPFD 처리구에서 25.6cm로 가장 길었으며, 엽폭은 처리 간 큰 차이는 없었으나 70% 차광 처리구에서 가장 넓었다. 이는 광 세기가 증가할수록 엽장이 짧아지는 경향을 보이며, 보광 수준에 따라 마디 수(엽 수) 형성이 차이를 나타내었다. 이러한 결과를 통해 보광 처리는 초장과 마디수가 증가하는 영향을 미쳤으나 엽장 및 엽폭 생장의 억제 효과를 나타낸 것으로 판단된다. 식물은 광량이 적을 때 광합성량을 증가시키기 위하여 엽면적을 넓히는데(Lusk, 2002; Haque 등, 2009) 엽면적은 30% 차광 처리구에서 14,890cm2·plant-1로 가장 크게 나타났으며, 70% 차광 처리구와 유의한 차이는 없었다. 그리고 보광 처리구는 작은 엽면적을 보였는데 이는 보광 환경에서 충분한 순광합성량에 도달하는 정도의 엽면적 크기에 도달했기 때문이라고 판단된다. 생체중 또한 30% 차광 처리구에서 가장 높았고, 두 보광 처리구는 상대적으로 낮았다. 그리고 건물중은 모든 처리구에서 유의한 차이가 나타나지 않았으며, 70% 차광 처리구에서 유의하게 낮았다. 과실 무게는 NL + SL 240PPFD 처리구에서 가장 무거운 3.0kg·fruit-1를 나타내었으나 70% 차광 처리구에서만 유의하게 가벼웠고, 다른 처리 간에는 유의한 차이가 없었다. 기존 수박 재배 방식인 포복재배보다 수직형 재배 시 상부 엽의 광합성량보다 하부의 광합성량이 그림자로 인해 감소하므로 과실 무게의 증가를 위한 충분한 엽면적 확보가 필요하다(Watanabe 등, 2001; Watanabe, 2014)는 결과를 통해 수직형 재배를 진행하였던 본 연구에서 NL + SL 240PPFD 처리구에서 엽면적은 가장 작았지만 마디 수(엽 수)가 많아 과실 비대를 위한 광합성량이 충분했던 것으로 생각된다. 그리고 모든 처리구에서 당도는 9.0-9.7Brix 범위로 통계적으로 유의한 차이는 없었지만 NL + SL 240PPFD 처리구에서 9.7Brix로 가장 높았다.
Table 1.
Comparison of growth parameters of seedless watermelon under shading and supplemental lighting treatments.
| Light levelsz |
Plant height (cm) |
No. nodes (ea·plant-1) |
Leaf length (cm) |
Leaf width (cm) |
| 70% Shading of NL | 214.5 cy | 26.3 b | 25.2 a | 26.4 a |
| 30% Shading of NL | 196.9 d | 21.5 c | 23.9 b | 25.0 b |
| NL | 241.5 b | 26.6 b | 23.9 ab | 25.2 bc |
| NL + SL 120PPFD | 254.6 ab | 23.8 b | 25.6 a | 26.3 a |
| NL + SL 240PPFD | 254.8 a | 29.2 a | 22.9 c | 23.5 c |
Table 2.
Comparison of leaf area, fresh weight, dry weight, fruit weight, and soluble solids content of seedless watermelon under different light levels
| Light levelsz |
Leaf area (cm2·plant-1) |
Fresh weight (g·plant-1) |
Dry weight (g·plant-1) |
Fruit weight (kg·fruit-1) |
Soluble solids content (Brix) |
| 70% Shading of NL | 14,016 ay | 1,102 a | 96 b | 2.3 b | 9.5 a |
| 30% Shading of NL | 14,890 a | 1,197 a | 129 a | 2.8 a | 9.0 b |
| NL | 12,392 ab | 1,109 a | 112 a | 2.5 a | 9.5 a |
| NL + SL 120PPFD | 8,659 b | 679 b | 117 a | 2.6 a | 9.5 a |
| NL + SL 240PPFD | 9,672 b | 792 b | 112 a | 3.0 a | 9.7 a |
본 연구에서는 광 수준에 따라 수박 식물체의 생장 및 과실 특성의 차이가 나타남을 확인할 수 있었으며, 온실에서 씨 없는 수박 재배 시 광·온도 환경의 복합적 관리가 단순히 생육 안정성뿐만 아니라 과실 품질 및 생산량 증가에 영향을 미치는 것으로 나타났다. 광 환경은 암꽃 발생과 과실 비대에 직접적으로 작용하였으며, 차광은 고온기 생육 스트레스 보완, 보광은 부족한 광합성 보완에 효과적이었다. 따라서 실제 재배 현장에서는 계절과 기상 조건에 따라 차광과 보광을 병행하는 정밀한 환경 제어 전략이 요구된다. 본 연구는 씨 없는 수박의 안정적 생산과 품질 확보를 위한 광·온도 관리 모델 구축의 기초 자료로 활용될 수 있으며, 향후 재배기술 개발에도 기여할 것으로 기대된다.
