

서 론
꽃은 식물의 영양생장 단계에서 축적된 에너지의 총체로, 생식생장의 개시를 의미한다(Lee et al. 2014). 최근 연구에 따르면, 꽃은 과채류나 엽채류처럼 항산화 성분을 풍부하게 함유하고 있어 기능성 식품 소재로 주목받고 있다(Park and Kim 2017). 이에 따라 식용 꽃에 대한 수요가 증가하고 있으며, 섭취 방법 또한 다양화되고 있다(Yoo et al. 2021). 특히 초롱꽃목 국화과 천수국속에 속하는 프렌치매리골드(Tagetes patula L.)는 처음에는 관상용 화훼로 도입되었으나(Kim et al. 2021), 최근에는 루테인과 플라보노이드 등 기능성 물질이 다량 함유되어 있어 건강식품 원료로의 가치도 재조명되고 있다(Kishimoto et al. 2005; Meda et al. 2005; Muley et al. 2009; Siriamornpun et al. 2012; Bhave et al. 2020). 매리골드에 함유된 루테인은 총 카로티노이드의 약 91%를 차지하며, 특히 눈 건강과 관련된 기능성으로 잘 알려져 있다.
이러한 기능성에도 불구하고, 매리골드를 비롯한 많은 화훼류는 개화 시기의 불확실성과 생육 예측의 어려움으로 인해 상업적 재배나 품질 관리에 어려움을 겪고 있다. 일반적으로 개화는 환경 스트레스에 반응하는 생리적 전환점이며, 광주기(photoperiod), 온도, 영양상태 등 다양한 환경 요인의 영향을 받는다(Thomas and Vince-Prue 1997). 하지만 광주기 반응은 개화 시점을 조절하는 요소 중 하나일 뿐, 생존을 좌우하는 절대적 요소는 아니다.
최근 스마트팜과 정밀농업 기술의 발전에 힘입어, 작물의 생육 상태를 자동으로 분석하고 예측하는 기술이 농업 분야에서 빠르게 도입되고 있다. 화훼류의 경우 개화는 상품성 판단, 수확 시기 조절, 생육환경 평가 등의 중요한 지표로 활용되기 때문에, 개화 시기의 자동 예측 및 모니터링 기술에 대한 수요가 증가하고 있다(Jiao et al. 2022). 그러나 여전히 많은 재배 현장에서는 숙련자의 경험과 시각적 판단에 의존하는 경우가 많고, 이는 주관성과 인력 의존성이라는 한계를 지닌다(Mann et al. 2022).
최근 딥러닝 기술의 발전과 함께 영상 기반 생육 분석 연구가 농업 분야 전반으로 확산되고 있다. 특히 합성곱 신경망(Convolutional Neural Network, CNN)은 이미지로부터 공간적 특징을 자동으로 추출하는 데 탁월한 성능을 보이며, 작물 생육 단계 분류, 병해 진단, 생체량 추정 등(Ghosal et al. 2018), 다양한 농업 영상 분석 분야에서 핵심적인 방법으로 활용되고 있다. CNN은 식물 영상에서 잎의 형태, 색상 변화, 꽃의 구조적 발달과 같은 시각적 정보를 정량화하는 데 널리 사용되어 왔다. 그러나 CNN은 기본적으로 단일 시점의 정적인 이미지 정보를 입력으로 처리하는 구조를 가지므로, 시간에 따라 점진적으로 변화하는 생육 과정이나 발달 단계를 충분히 반영하는 데에는 한계가 있다. 따라서 단일 이미지 기반 CNN만으로는 개화율이 유사한 시점 간의 미세한 차이를 구분하거나, 향후 개화 진행 정도를 예측하는 데 제한이 발생할 수 있다. 이러한 한계를 보완하기 위한 방법으로 순환 신경망(Recurrent Neural Network, RNN)이 제안되어 왔다. RNN은 이전 시점의 정보를 내부 상태(hidden state)에 저장하여 현재 출력에 반영할 수 있는 구조를 가지며, 시계열 데이터 분석에 적합한 딥러닝 모델로 알려져 있다. 이러한 특성으로 인해 RNN은 생육 환경 데이터, 생장 곡선, 생산량 예측과 같은 농업 시계열 분석 연구에서도 점차 활용 범위가 확대되고 있다. 최근에는 CNN과 RNN을 결합한 하이브리드 구조가 제안되어, 영상의 공간적 특징과 시간적 변화를 동시에 고려하는 연구들이 보고되고 있다. CNN을 통해 각 시점의 이미지로부터 공간적 특징을 추출한 뒤, 이를 시계열 형태로 RNN에 입력함으로써 시간에 따른 형태 변화와 발달 경향을 학습하는 방식이다. 이러한 CNN+RNN 구조는 단일 이미지 기반 분석보다 생육의 연속성을 효과적으로 반영할 수 있어, 개화 시점 예측, 생육 단계 추정, 작물 발달 곡선 모델링 등에 적합한 접근법으로 평가되고 있다(Kamilaris and Prenafeta-Boldú, 2018).
매리골드는 생육 기간이 짧고 개화 과정이 명확하게 구분되는 대표적인 화훼류로, 시계열 기반 개화 예측 연구의 대상으로 적합하다. 본 연구에서는 시간 흐름에 따라 수집된 연속 이미지를 활용하여 개화 진행률을 예측하는 CNN+RNN 하이브리드 모델을 제안하였다. 본 모델은 기존 정적 이미지 기반 모델보다 시간 정보를 반영함으로써 더 정밀한 개화 추적이 가능하며, 향후 다양한 화훼 작물의 생육 모니터링 자동화에 기여할 수 있는 기술적 기반을 제공할 수 있을 것으로 기대된다.
재료 및 방법
1. 실험 재료
프렌치매리골드(Tagetes patula L.) ‘Durango Red’ 종자(Hanmi Seeds Co., Chungju, Korea)를 우레탄 스폰지(3.5× 3.5×3.0cm)에 파종하여 사용하였다. 실험은 완전제어형 식물공장에서 수경재배로 진행되었으며, 온도는 20-22℃, 상대습도는 55-60%로 설정하였다. 명기/암기 12/12 광주기 하에서, 박막수경재배방식(nutrient film technique, NFT)으로 재배하였다. Hoagland 배양액(Hoagland and Arnon 1950)을 본엽 1쌍(2매)이 전개되었을 때부터 공급하였고, 배양액의 pH와 EC는 각각 5.9-6.1, 1.5dS·m-1로 관리하였다. 꽃봉오리의 평균 지름이 약 0.5cm일 때부터 측정을 시작하였으며(Fig. 1B), 측정 간격은 주간 1시간, 야간 3-5시간 간격이었다.
2. 데이터셋 구성
개화 초기부터 완전히 꽃이 핀 시점까지 약 1시간 간격으로 정면 이미지(iPhone 12 Pro, Apple Inc., Cupertino, CA, USA)를 촬영하였으며, 개별 식물체마다 하나의 폴더로 분류하여 저장하였다. 이미지 파일명은 촬영 시점을 시간 단위로 명명하였으며(예: 0.jpg, 5.jpg, ..., 72.jpg), 폴더명은 식물체 식별번호로 구성하였다. 개화단계 사진은 Fig. 1과 같다.
이미지 해상도는 1440×1440 픽셀이며, 학습 효율성을 위해 전처리 단계에서 128×128 크기로 리사이즈 하였다. 각 시점의 개화 진행률은 현재 시간/최대 시간으로 계산하여 0에서 1 사이의 실수로 정규화하였으며, 이는 회귀 모델의 타깃 레이블로 사용되었다.

3. 시계열 학습을 위한 데이터 구성
영상의 시계열성을 반영하기 위해 연속된 이미지 5장을 하나의 입력 시퀀스로 구성하였다. 각 시퀀스는 시간 순서대로 정렬된 5장의 이미지로 구성되며, 마지막 이미지 시점의 개화 진행률을 예측 대상으로 설정하였다. 전체 데이터 중 80%는 학습용으로, 나머지 20%는 검증용으로 무작위 분할하여 사용하였다. 참고적으로, 본 연구에서 구성된 시계열 입력 시퀀스는 동일한 식물 개체를 시간에 따라 연속적으로 추적하여 촬영한 이미지들로 구성되었으며, 각 시퀀스는 개체 단위의 시간적 변화만을 반영하도록 설계되었다. 데이터 분할 과정에서는 동일 식물 개체에서 생성된 시퀀스가 학습용과 검증용 데이터셋에 동시에 포함되지 않도록 개체(plant individual) 단위로 분리하여 무작위 분할을 수행하였다.
4. 모델 구조
제안된 CNN+RNN 시계열 모델의 성능을 정량적으로 평가하기 위하여, 동일한 데이터셋을 기반으로 다양한 딥러닝 구조를 비교 모델로 설정하고 동일한 조건에서 학습 및 검증을 수행하였다. 비교 대상으로는 일반적인 합성곱 기반 회귀 모델인 BasicCNN을 포함하여, 대표적인 사전학습(pretrained) 백본 구조인 ResNet18(He et al. 2016), MobileNetV2(Sandler et al. 2018), EfficientNetB0(Tan and Le 2019)를 선정하였다. 비교 모델은 시계열 정보를 고려하지 않고, 각 시점의 단일 이미지를 입력으로 사용하여 개화 진행률을 예측하도록 구성되었다. 각 모델의 마지막 출력층은 회귀 문제에 맞추어 단일 뉴런과 Sigmoid 활성화 함수를 적용하였다. 사전학습된 백본을 사용하는 모델은 ImageNet 데이터셋으로 학습된 가중치를 초기값으로 적용하고, 전이학습(fine-tuning)을 통해 전체 구조를 학습하였다. 학습 조건은 제안 모델과 동일하게 설정하였으며, 학습률은 0.001, 옵티마이저는 Adam, 손실함수는 평균제곱오차(MSE)를 사용하였다. 각 모델은 에폭스100동안 학습하였으며, 동일한 검증 데이터셋을 기반으로 평균절대오차(MAE), 결정계수(R2) 등의 성능 지표를 계산하였다. 이를 통해 시계열 정보를 반영하지 않은 기존 구조들과 제안 모델 간의 성능 차이를 비교 분석하였다.
제안된 모델은 CNN과 RNN을 결합한 시계열 딥러닝 모델로, 전체 구조는 다음과 같다.
1) CNN 특징 추출기
각 이미지에 대해 동일한 합성곱 신경망(CNN) 모듈을 적용하여 시각적 특징을 추출하였다. 이 모듈은 3개의 합성곱 층과 최대 풀링 층으로 구성되며, 마지막 단계에서 평균 풀링을 통해 1차원 벡터(64차원)를 출력한다.
2) LSTM 기반 RNN 모듈
CNN에서 추출된 시계열 벡터들을 입력으로 받아 시간 축에 따라 학습을 수행한다. 은닉 상태 차원은 128로 설정하였다.
3) 회귀 출력층
LSTM의 마지막 타임스텝 출력을 입력으로 하여 전결합층을 통해 0-1 범위의 개화 진행률을 예측한다. 출력층에는 Sigmoid 활성화 함수를 적용하여 정규화된 값을 도출하였다.
5. 모델 학습 및 구현 환경
모델은 PyTorch(버전 1.13)를 기반으로 구현되었으며, Adam 옵티마이저(학습률 0.001)를 사용하여 평균제곱오차(MSE)를 최소화하도록 학습하였다. 전체 학습 에폭스(epoch)은 100으로 설정하였으며, 배치 크기는 16이다. 학습은 NVIDIA GPU 환경(GeForce RTX 3090)에서 수행되었다.
6. 성능 평가
예측 성능 평가는 검증 데이터셋을 대상으로 다음과 같은 세 가지 지표를 사용하였다:
•MSE (Mean Squared Error) = : 평균제곱오차
•MAE (Mean Absolute Error) = : 평균절대오차
•R2 (Coefficient of Determination) =
: 결정계수, 회귀 성능을 총변동 대비 예측력으로 표현
여기서 yi는 실제 개화 진행률, ŷi는 모델의 예측값, ӯ는 실제값의 평균, n은 데이터 개수를 의미한다.
또한, 예측 결과와 실제 레이블 간의 산점도를 시각화하여 예측 일치도를 확인하였다. 이를 통해 제안된 모델이 개화 진행 과정을 정량적으로 얼마나 정확하게 추정하는지를 분석하였다.
결과 및 고찰
본 연구에서 제안한 CNN+RNN 모델은 MSE 0.0006, MAE 0.0196, R2 0.993을 기록하며, 비교 모델들에 비해 관측값에 대한 매우 높은 적합도를 보였으며, 예측 오차 또한 현저히 낮은 수준으로 나타났다(Table 1). 특히 R2 값이 0.99 이상으로 나타난 점은, 모델이 실제 개화 진행률의 분산을 거의 완전히 설명할 수 있음을 의미하며, 회귀 모델의 성능 척도 중 가장 신뢰도 높은 지표로 평가된다. 이러한 결과는 시계열 정보를 반영하는 모델이 정적인 이미지 기반 예측보다 실제 생육 변화를 더 효과적으로 학습할 수 있음을 시사한다.
Table 1
Comparison of blooming progress prediction performance among deep learning models
| Model | MSEz | MAEy | R2x |
|
BasicCNN ResNet18 MobileNetV2 EfficientNetB0 CNN+RNN |
0.0145 0.0966 0.0966 0.0914 0.0006 |
0.0972 0.2711 0.2709 0.2599 0.0196 |
0.849 -0.006 -0.006 0.048 0.993 |
반면, ResNet18, MobileNetV2, EfficientNetB0 등의 사전학습(pretrained) 기반 모델들은 높은 표현력을 갖추고 있음에도 불구하고 본 실험에서는 오히려 MSE가 약 0.091 내외로 높고, R2 또한 0에 가깝거나 음수(-0.0053까지)로 나타났다. 이는 모델이 데이터 분산을 제대로 설명하지 못했음을 의미하며, 실제로 과적합(overfitting) 또는 시계열 특성 반영의 한계로 인해 일반화 성능이 저하되었음을 나타낸다(Ma et al. 2024). 특히 단일 이미지로만 판단하는 기존 CNN 구조는 개화율이 비슷한 이미지(예: 40%와 60%) 간의 차이를 구분하는 데 어려움을 겪을 수 있다.
예측값과 실제값의 관계를 시각화한 산점도(Fig. 2)를 보면, CNN+RNN 모델의 예측 결과는 대부분 y = x의 대각선 근처에 밀집되어 있어, 예측값과 실제값 간의 편차가 작고 정확도가 높다는 것을 직관적으로 확인할 수 있다. 이와 같은 결과는, 모델이 개화 진행 과정에서의 미세한 시각적 변화를 누적적으로 인식하고 반영할 수 있었음을 보여준다. CNN+RNN 구조는 각 이미지에서 시각 정보를 추출한 후, 시간 순서를 반영하는 LSTM 구조를 통해 시계열 패턴을 학습함으로써 생육 변화의 맥락을 효과적으로 파악하였다.
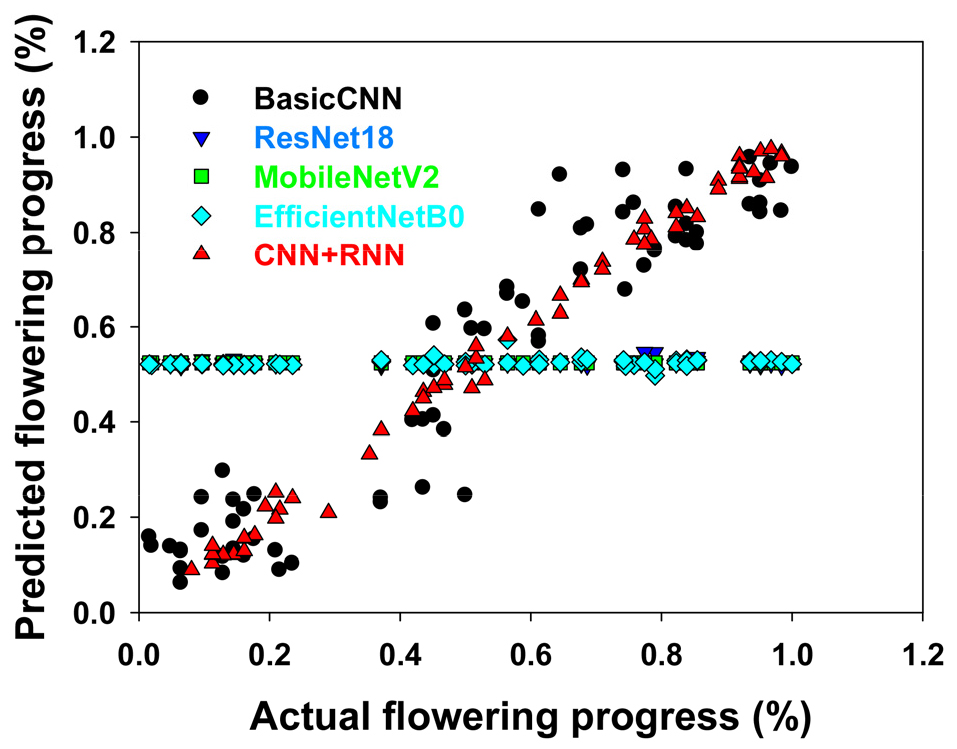
개화는 단일 시점의 정적인 상태라기보다는 생육 축을 따라 점진적으로 누적된 구조적 및 색상적 변화의 총합으로 이해될 수 있으며(Ghosal et al. 2018; Singh et al. 2020), 이러한 시계열적 특성은 전통적인 CNN 구조로는 포착하기 어렵다(Lv et al. 2019). 제안된 모델은 이러한 누적 변화를 학습할 수 있도록 구성되어, 정적인 이미지 하나만을 기준으로 판단하는 방식보다 훨씬 정교한 개화율 예측이 가능하였다. 실제로도 CNN+RNN 모델은 단일 시점에서 개화율을 정확히 구분하기 어려운 유사 이미지 간에도 시간 흐름에 따른 변화를 활용하여 오차를 줄이는 데 기여하였다.
이러한 결과는 영상 기반 생육 분석에서 시계열 정보를 고려한 접근 방식이 예측 성능 향상에 기여할 수 있음을 보여준다. 특히 화훼 작물의 경우 잎의 확장, 꽃봉오리의 팽창, 꽃잎의 전개 등 개화에 이르는 외형 변화가 시간 흐름에 따라 단계적으로 발생하는 특성을 가지므로(Lee et al. 2014), 시계열 모델이 개화 진행 과정을 보다 정밀하게 반영하는 데 적합한 구조임을 확인할 수 있었다.
또한, 제안된 CNN+RNN 구조는 제한된 데이터셋으로도 높은 예측 성능을 달성하였다는 점에서 실용성 측면에서도 의미가 있다. 실제 농업 현장에서는 대규모 학습 데이터 확보가 어려운 경우가 많기 때문에, 소규모 데이터셋에서도 높은 성능을 보장하는 모델의 필요성이 크다(Yu et al. 2022). 본 모델은 그와 같은 조건에서도 유의미한 성과를 보여, 적용성 측면에서 경쟁력이 높다고 판단된다. 다만, 매리골드는 다화성 식물로서 하나의 개체에서 여러 개의 꽃봉오리가 시간차를 두고 지속적으로 발달하는 생육 특성을 가진다. 본 연구에서는 시계열 영상 기반 개화 진행률 예측 모델의 개념적 타당성과 구조적 유효성을 검증하는 것을 주된 목적으로 하였으며, 이에 따라 정면에서 촬영된 단일 꽃봉오리를 대상으로 개화 과정을 추적·분석하였다. 따라서 동일 개체 내 다수의 꽃봉오리가 동시에 존재하거나, 잎이나 줄기에 의해 일부 꽃봉오리가 가려지는 실제 재배 환경의 복잡성은 본 연구의 분석 범위에 포함되지 않았다. 그럼에도 불구하고, 본 연구에서 제안한 CNN+ RNN 기반 시계열 모델은 단일 꽃봉오리의 시간적 개화 패턴을 정량적으로 학습하고 예측할 수 있음을 실험적으로 입증하였으며, 이는 향후 개체 단위 개화율 산출 기술로 확장될 수 있는 기초 기술로 활용 가능하다고 판단되었다. 예를 들어, 객체 탐지 기반 모델(YOLO 등)을 활용하여 개체 내 다수의 꽃봉오리를 자동으로 검출·추적한 후, 각 꽃봉오리에 대해 본 연구의 시계열 개화 예측 모델을 개별적으로 적용함으로써, 개체 전체의 평균 개화율이나 개화 분포를 정량적으로 산출하는 방식으로의 확장이 가능할 것으로 판단된다.
향후 연구에서는 다음과 같은 방향으로 확장이 가능할 것으로 기대된다. 첫째, 다양한 품종과 환경 조건(온도, 광주기, 습도 등)을 반영한 일반화 성능 검증이 필요하다. 둘째, 더 긴 시계열 시퀀스(예: 10장 이상)를 활용한 예측 정확도 향상을 도모할 수 있으며, Transformer 기반의 구조를 추가적으로 비교하는 것도 고려할 수 있다(Liu et al. 2021). 셋째, 본 연구에서는 개화율만을 정량화하였으나, 이후 생장속도 추정, 최대 개화 지점 탐지, 낙화 시점 예측 등의 응용 분야로도 확장 가능하다. 넷째, UAV 기반 촬영 영상, 멀티스펙트럼/하이퍼스펙트럼 영상, 열화상 등 다양한 영상 센서를 통합하는 융합 모델 연구를 통해 정밀 농업의 실제 활용도 제고가 가능하다(Sankaran et al. 2015). 또한 본 연구에서 제안한 CNN+RNN 기반 시계열 모델은 연속된 5장의 이미지를 입력으로 사용한 반면, 비교군으로 설정한 기존 CNN 기반 모델들은 단일 이미지를 입력으로 사용하였다. 이러한 입력 구성의 차이는 제안 모델의 성능 향상이 순수하게 시계열 구조의 효과에 기인한 것인지, 혹은 입력 정보량 증가에 따른 영향이 일부 포함된 것인지에 대한 추가적인 검증의 필요성이 있다. 즉, 동일한 CNN 백본 구조에 대해 다중 이미지 입력을 적용하거나, 시퀀스 내 이미지의 시간적 순서를 교란하는 ablation study를 수행함으로써, 제안 모델이 시간적 연속성에 기반한 형태 변화 자체를 학습하고 있는지 여부를 보다 명확히 확인할 수 있을 것이다. 이러한 추가 분석은 시계열 정보의 기여도를 정량적으로 분리·평가하는 데 기여할 뿐만 아니라, 본 연구에서 제안한 접근법의 일반화 가능성과 해석 가능성을 한층 강화할 수 있을 것으로 판단된다. 향후 연구에서는 이러한 보완 실험을 통해 모델 구조와 입력 구성 간의 영향을 체계적으로 분석하고, 다양한 화훼 및 작물 생육 예측 문제로의 확장 가능성을 검증하고자 한다.
결론적으로 본 연구는 매리골드를 대상으로 시계열 영상 기반 개화 예측 모델을 구축함으로써, 화훼 작물의 생육 분석 자동화에 기술적 기초를 제공하였다. 제안된 CNN+RNN 구조는 시계열 정보를 효과적으로 학습하여 높은 정확도를 달성하였고, 향후 스마트팜, 자동화 생육 진단, 디지털 육종 등 다양한 농업 응용 분야에 적용 가능성이 클 것으로 판단된다.
