

Introduction
Abscisic acid (ABA) is an important signaling hormone which makes resistance and adaptation in plants against various abiotic stress conditions (Bakhsh et al., 2011; Li et al., 2010). ABA protects plants from water stress damage by inducing stomatal closure (Li et al., 2000) and increasing hydraulic conductance for water movement from roots to leaves (Ludewig et al., 1988; Zhang et al., 1995). Additionally, ABA has been reported to improve drought tolerance in various plant species, including Tradescantia virginiana L. (Frank and Farquhar, 2001), Pinusbanksiana L. (Rajasekaran and Blake, 1999), and Capsicum annuum L. (Leskovar and Cantliffe, 1992). Generally, ABA is regarded as an inhibitor of shoot and root growth of plants (Munns and Cramer, 1996). However, under certain conditions low concentrations of ABA applied either to intact roots, root segments, or excised roots growing in liquid culture has been found to stimulate the plant growth (Pilet, 1983; Pilet and Rebeaud, 1983; Takahashi et al., 1993). In addition, Pilet and Barlow (1987) reported that high or low concentrations of ABA can either inhibit or stimulate root growth, respectively.
The importance of ABA in stress regulation is widely recognized for a long time. While ABA can be produced in the roots of many plants and is transferred through the xylem to the leaves when the plant is exposed to low moisture conditions (Jiang and Hartung, 2008), information about the effect of the application of ABA in nutrient solution on growth and physiology and alleviation in drought stress of tomato seedlings is limited. Therefore, the objective of this study was to examine effect of shortterm ABA application to the nutrient solution on the maintaining of the quality of tomato seedlings in water stress condition.
Materials and Methods
1. Plant materials and growing conditions
Seeds of ‘Dotaerang Dia’ tomato were sown in the 128- cell plug trays filled with commercial growing substrate (BM2, Berger Group Ltd, Canada). Twenty-five days after sowing, the seedlings were transplanted to a hydroponic( NFT) system in a plastic house at Kangwon National University from June to July 2014. The nutrient solution was adjusted to EC 1.5dS·m–1 by using 1g·L–1 of ‘Wonder Grow’(Chobi Co., Ltd., Korea) fertilizer, consisting percentage of N: P: K: Mg: B: S: Mn: Fe: Zn such as 10: 8: 25: 2: 0.1: 5: 0.05: 0.05: 0.01, respectively.
2. ABA application and water stress treatment
Four ABA concentrations (0.5, 1, 2, and 3mg·L–1) and control (non-treatment) were applied to the nutrient solution in the DFT system (Fig. 3-A). On the 5th and 10th day after transplant to the hydroponic solution containing ABA with various concentrations, seedlings were transferred to - 5 bars of PEG-8000 (Sigma-Aldrich, Co., St. Louis, MO, USA) in a growth chamber (Hanbaek Co., Ltd., Bucheon, Korea) to induce water stress. The osmotic pressure was selected in various osmotic pressures (-1, -5, -10, -15 bars) of our pre- experiments (Data not showed). Sixty hours after treating in -5 bars of PEG solution, the seedlings were re-irrigated by transferring to the normal nutrient solution. Growth chamber conditions were as follows: relative humidity was maintained at 40-50%; light intensity was approximately 100 mmol·m–2·s–1 PFD provided by fluorescent lamps (Orex Co., Ltd., Goyang, Korea); temperature was set at 25°C. The osmotic pressure (OP) of PEG-8000 solution was calculated by equation (1) according to Michel (1983).

Where C = PEG concentration; T = Temperature (25°C).
3. Data collection and analysis
On the 10 days after transplant to the hydroponic solution with various ABA concentrations, the seedlings were evaluated for growth and physiological characteristics. The growth measurements included the seedling height (cm), number of leaves, leaf area (cm2) by leaf area meter (Area meter, Delta-T, UK), leaf chlorophyll content by using a chlorophyll meter (SPAD-502, Minolta, Japan), and fresh and dry weights of shoot and root. The dry weight of shoot and root were taken through oven-dry method at 80°C for 72 h until constant weight was achieved. The T/R ratio (shoot dry weight/root dry weight ratio) and compactness (shoot dry weight/plant height) were calculated according to Kim et al. (2008).
The root morphology such as total root surface area, total root length, average root diameter, and number of root tips was analyzed using Epson 10000XL scanner equipped with the WIN MAC RHIZO V 2009c program (Regent Instruments Inc., Canada) according to Arsenault et al. (1995). The roots were detached from their shoots and then placed in a tray (40 cm length × 30cm width × 2cm height) with water.
The physiological characteristics such as stomatal diffusive resistance, transpiration, and leaf temperature were assessed with an LI-1600 steady state porometer (LI-COR, Lincoln, Nebraska, USA) for the 4th leaf from the top of 5 plants of each treatment. Data were collected between 11.00-13.00 hrs.
The wilted or recovered seedlings were calculated when 75% of leaves per seedling withered or recovered, respectively.
The experiments were arranged in completely randomized design. For the statistical analysis of growth and physiological parameters, six seedlings per treatment were randomly selected. Data were analyzed using SASv.9.3 software (SAS Institute Inc., Cary, NC, USA). Mean separations were calculated using Duncan’s multiple range test at P ≤ 0.05.
Results and Discussion
1. The effect of short-term application of ABA to nutrient solution on growth characteristics of tomato seedlings
The changes of the plant growth characteristics to ABA treatment are given in Table 1,2 and Fig. 1. Plant height and stem diameter of tomato seedlings were significantly decreased with ABA concentration. These results agree with previous reports that ABA acts as an inhibitor of shoot growth (Munns and Cramer, 1996). In this study, the tallest plant height and thicker stem diameter were observed in the control, but for the plant height the difference was not statistically significant among the control, 0.5, and 1mg·L–1 ABA treatments. For the stem diameter, no statistical differences were observed in the 0.5, 1, and 2mg·L–1 treatments, but all were lower than the control. The height and stem diameter decreased at 2 and 3mg·L–1 ABA concentration.
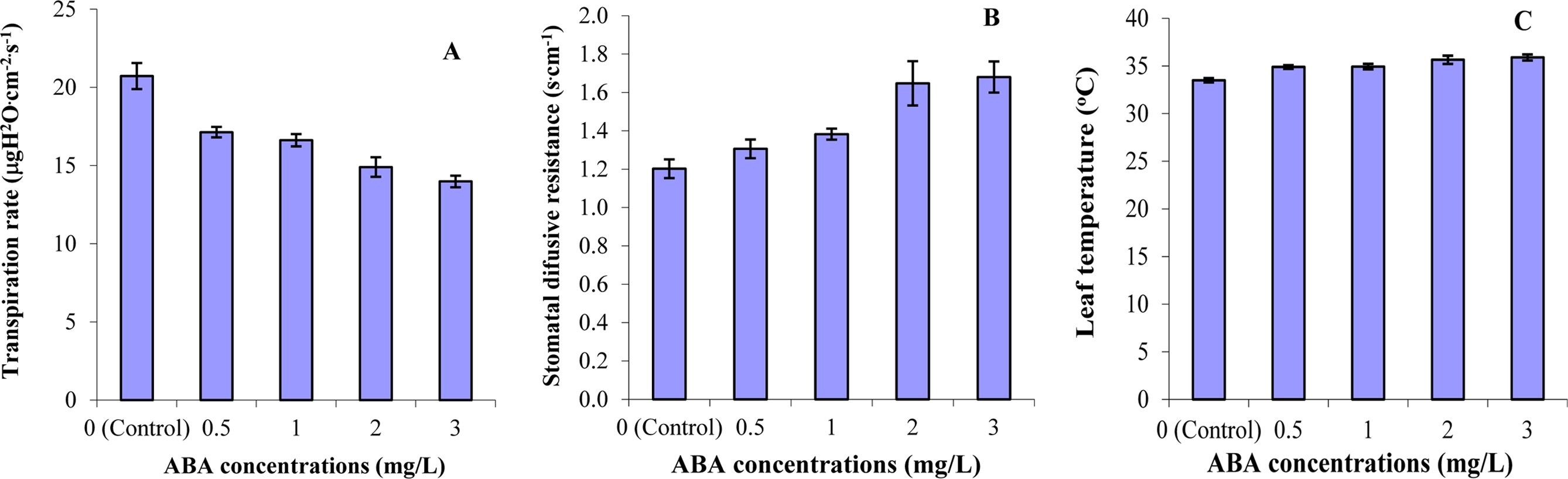
Table 1.
Effect of ABA concentrations in nutrient solution on growth characteristics of tomato seedlings at 10 days after treatment.
| ABA (mg/L) | Plant height (cm) | Stem diameter (mm) | No. of leaves | Leaf length (cm) | Leaf width (cm) | Leaf chlorophyll value (SPAD) | Leaf area (cm2) |
|---|---|---|---|---|---|---|---|
| 0 (control) | 20.38az | 7.85 a | 8.00 a | 22.70 a | 16.15 a | 41.31 a | 335.33 a |
| 0.5 | 19.31 a | 7.08 b | 7.63 ab | 20.51 ab | 14.59 ab | 41.29 a | 306.75 a |
| 1 | 19.13 a | 7.14 b | 8.00 a | 21.00 ab | 15.11 ab | 40.18 a | 311.75 a |
| 2 | 17.31 b | 6.93 b | 7.25 b | 20.65 ab | 14.09 b | 39.85 a | 240.75 b |
| 3 | 17.25 b | 6.40 c | 7.00 b | 19.25 b | 12.74 c | 39.50 a | 216.75 b |
The inhibition of leaf growth by ABA was reported in many papers (Alves and Setter, 2000; Carrow, 1996; Sharp et al., 1994). Thus not surprisingly in this study, the leaf number, leaf length, leaf width, and leaf area decreased with increasing ABA concentration. The leaf number and leaf area, however, were similar in the control, 0.5, and 1mg·L–1, but significantly decreased in the seedlings treated with 2 and 3mg·L–1 of ABA. There was no statistically significant difference observed in the leaf length among control, 0.5, 1, and 2mg·L–1 of ABA, but a significant decrease was found in the 3mg·L–1 treatment. The leaf width was similar in seedlings treated with 0.5, 1, and 2mg·L–1 of ABA, but lower than that in the control. The lowest value of leaf width was observed in seedlings treated at 3mg·L–1 of ABA. Leaf chlorophyll value decreased with increasing ABA concentration. The highest value of chlorophyll content was observed in seedlings in the control and the lowest value was observed in the 3mg·L–1 treatment. These results agree with Farooq and Bano, (2006) and Iqbal et al. (2010) who observed that ABA application causes decreases in chlorophyll contents in wheat, mungbean, and chickpea plants. However, in this study the differences in the chlorophyll values were not statistically different in all treatments (Table 1).
ABA added to the nutrient solution of intact plants causes a greater inhibition of shoot growth than root growth (Creelman et al., 1990; Robertson et al., 1990), and sometimes it causes a sustained increase in root growth (Biddington and Dearman, 1982; Watts et al., 1981). In this study, the fresh and dry weight of shoot decreased with increasing ABA concentration. However, there were no statistical differences between the control, 0.5, and 1mg·L–1 of ABA treatments, but it decreased with ABA concentration of 2 and 3mg·L–1. The highest value of fresh and dry weight of root was observed in the 1mg·L–1 of ABA treatment (Table 2). The root weight increase may be caused indirectly by ABA inhibiting shoot growth, thus resulting in a redistribution of assimilates to the roots. Alternatively, ABA may actively promote root growth and in so doing direct the movement of metabolites away from the growing point of the shoot to the root. These results agree with Abou-Mandour and Hartung (1980) who also shown that ABA increases the weight of adventitious roots on the hypocotyls of Phaseoluscoccineus seedlings. The seedlings treated with 0.5 and 1mg·L–1 ABA were similarly compact as in the control, but the compactness decreased in the 2 and 3mg·L–1 treatments. The T/R ratio decreased with ABA concentration (Table 2), similarly as reported for the root to shoot dry weight ratio of Capsicum annuum and Zea mays (Watts et al., 1981).
Table 2.
Effect of ABA concentrations in nutrient solution on fresh and dry weight of shoot and root, T/R, and compactness of tomato seedlings at 10 days after treatment.
| ABA concentration (mg/L) | Fresh weight (g) | Dry weight (g) | T/R ratioz | Compactnessy (mg/cm) | ||
|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | |||
| 0 (control) | 26.41 ax | 3.91 b | 1.98 a | 0.26 b | 7.62 a | 9.71 a |
| 0.5 | 25.74 a | 4.08 b | 1.80 a | 0.25 b | 7.34 a | 9.32 a |
| 1 | 24.72 a | 4.89 a | 1.85 a | 0.30 a | 6.14 b | 9.65 a |
| 2 | 17.95 b | 4.14 b | 1.34 b | 0.23 c | 5.91 b | 7.75 b |
| 3 | 16.64 b | 3.77 c | 1.17 c | 0.22 c | 5.63 b | 6.80 b |
The transpiration rate decreased significantly with increasing ABA concentration, while the stomatal diffusive resistance and leaf temperature increased (Fig. 1), which agrees that ABA induces stomatal closure, resulting in decreased transpiration (Arteca et al., 1985; Mansfield and Jones, 1971; Mittelheuser and Van Steveninck, 1969). Jones and Mansfield (1972) also showed that ABA reduces transpiration in barley (Hordeumvulgare L.). Moreover, Takahashi et al. (1993) reported that the leaf temperature of upper, middle, and lower leaves of tomato plants increased with increasing ABA concentration in the culture solution.
2. The effect of short-term application of ABA to nutrient solution on root morphology of tomato seedlings
The changes in root morphology in response to ABA treatment are summarized in Fig. 2. The total root surface area, total root length, average root diameter, and number of toot tips were significantly different among the various ABA treatments. Compared with control, the total root surface area increased at 0.5, 1, and 2mg·L–1 of ABA, but decreased in seedlings treated with 3mg·L–1 of ABA. The highest value of total root surface area was observed at 1mg·L–1 of ABA. The total root length increased at 0.5 and 1mg·L–1 of ABA, but decreased at 3mg·L–1 of ABA. The decrease in the total root length may be caused directly by ABA inhibition of the root development. These results agree with Liao et al. (2008) who found that ABA exposure at low concentration (0.5μM) increased the root length and at higher concentrations (1 and 5μM) the root length decreased in mutant soybean plants. Moreover, it has been previously shown that ABA does not increase the length of the roots (Gaither et al., 1975; Watts et al., 1981; Yamaguchi and Street, 1977). The highest value of the average root diameter was observed at 1mg·L–1 of ABA, while it was similar in the control or 0.5, 2, and 3mg·L–1 of ABA. The number of root tips increased with ABA treatment, but decreased with high concentration (Fig. 2), which is similar with the results of Biddington and Dearman (1982) who reported that the most obvious effect of ABA on cauliflower root growth was to increase the number of lateral branches and to increase both the number and length of the root hair.
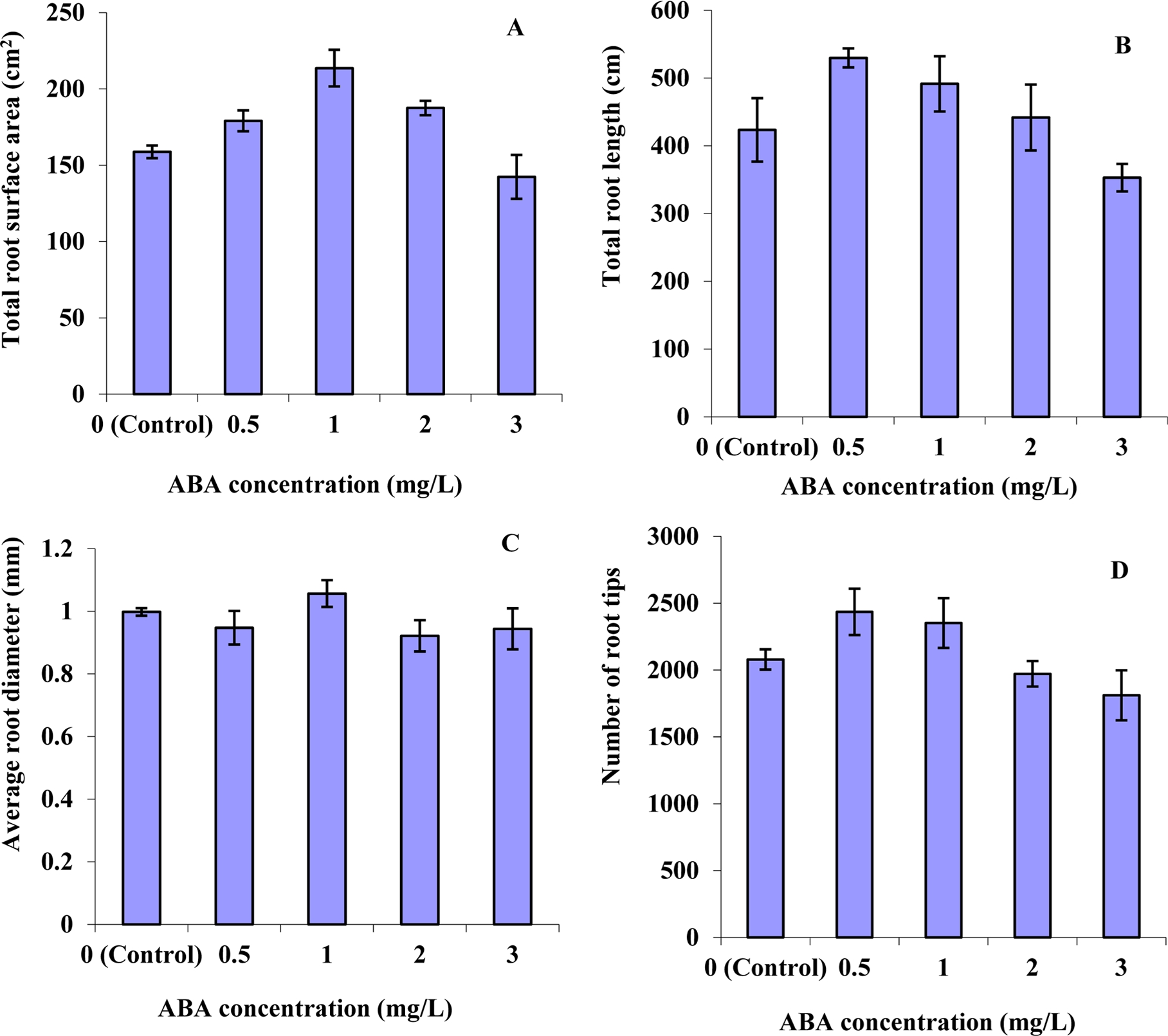
Fig. 2.
Effect of ABA concentrations in nutrient solution on root morphology of tomato seedlings at 10 days after treatment. (A) Total root surface area (cm2), (B) total root length (cm), (C) average root diameter (mm), and (D) number of root tips of tomato seedlings. Vertical error bars represent standard errors (n=6).
3. The effect of short-term application of ABA to nutrient solution on drought stress of tomato seedlings
ABA accumulation protects plants from drought damage by inducing stomata closure to reduce water loss via transpiration (Borel et al., 1997; Li et al., 2000) and increasing hydraulic conductance for water movement from roots to leaves (Ludewig et al., 1988; Zhang et al., 1995). In this study, the application of ABA in nutrient solution enhanced the drought tolerance of tomato seedlings by delaying the starting time of wilting point under drought conditions and enhancing the recovery after re-irrigation. The starting time of wilting point was observed at 10 hrs in the control treatment, but it was observed at 30 hrs in all ABA treatments. The 100% of wilted plant was observed in the control treatment after 20 hrs of water stress, but it was observed at 40 hrs in treated seedlings with 2 and 3mg·L–1 of ABA for 5 days and at 50 hrs in other ABA treatments. The seedlings grown in nutrient solution with ABA for 5 days, 83.3% and 66.7% of surviving seedlings were found in the 0.5 and 1mg·L–1 treatments after re-irrigation, while no surviving seedlings were found in the control, 2 and 3mg·L–1 of ABA treatment. In the seedlings grown in nutrient solution with ABA for 10 days, the highest recovery of the seedlings post re-irrigation was observed in the 0.5 and 1mg·L–1 ABA treatments. The seedlings in the 2 and 3mg·L–1 of ABA treatment had 66.7% and 50% survival seedlings,
respectively (Table 3). These results agree with those of Waterland et al. (2010) who reported that both spray and drench applications of 500mg·L–1 of ABA on a variety of popular bedding plants delayed the time to wilting as much as 3 days for marigolds and 4 days for petunia. Moreover, pansy maintained higher turgor when receiving ABA as a spray, whereas other crops, including petunia and impatiens, were more receptive to drench applications.
Table 3.
Effect of ABA concentrations in nutrient solution on drought tolerance of tomato seedlings grown in nutrient solution with ABA for 5 and 10 days.
In conclusions, the application of ABA in the nutrient solution enhanced the drought tolerance of tomato seedlings by delaying the start time wilting point and enhancing the recovery after re-irrigation. Short-term application of 1mg·L–1 of ABA in the nutrient solution stimulated the root growth and drought tolerance of tomato seedlings.
