

서 론
여름철 시설 채소재배를 위해 전일 차광(全日遮光) , 수시차광(隨時遮光), 자연 및 강제 환기, fog system 등이 이용되고 있으며, 최근에는 지열 히트펌프, 공기열 히트펌프에 의한 냉방 시스템이 개발되어 활용되고 있다. 그런데 대부분의 시설 농가는 온실 냉방을 위한 과다한 시설투자가 농가경영에 상당한 부담으로 작용하기 때문에 여름철 고온에 대한 적극적인 시설투자 보다는 휴작, 작기 조절 등 소극적인 재배적 방법으로 대처하고 있어 시설이용율 제고에서 많은 문제로 대두되고 있다(Woo 등, 1996).
여름철에 자연환기 만을 이용할 경우 온실 내부의 최고기온은 약 45℃에 달하며, 40∼50% 차광과 강제환기를 병행하더라도 과채류의 최고생육한계 기온인 35℃ 이하로 하강시키기에는 역부족이며, 물의 증발잠열능력을 이용한 fog system이 도입되어야만 30℃ 수준으로 하강시킬 수 있다(Woo 등, 1996). 그러나 이러한 물의 증발잠열 능력을 이용한 시스템은 야간에는 사용할 수 없으며, 다습한 지역에서는 시스템의 효과가 상당히 감소되어 시설의 효율성이 떨어진다. 또한, 시스템 설치비용이 많이 소요되므로 경제적 측면에서 농가에 부담이 될 수 있다. 따라서 최근에는 과다한 시설투자 없이 차광 및 강제 환기를 비롯한 기본적인 온도 하강장치(Woo 등, 1995; Woo 등, 2014)를 사용하거나 과산화수소(Woo 등, 2006), 기체 이산화탄소 시용(Woo 등, 2015), 액체 이산화탄소 엽면시비(Woo 등, 2019) 등으로 자체적으로 작물의 내서성 향상을 유도하는 기술이 개발되어 이용되고 있다.
여름철 온실 내부는 45℃ 이상의 극고온과 700W·m-2 이상의 강한 일사량으로 작물은 기공을 닫아 광 호흡을 하게 되고, 이는 과다한 증산작용을 억제하여 작물자체를 보호하기 위한 대사과정 중의 하나이다. 광 호흡은 C3 작물의 특징으로 한 여름 기온이 높고 일사 강도가 높을 때에 과다한 증산을 억제하기 위해 기공을 닫는 경우가 있다(Woo 등, 2014). 그 결과 잎 내부의 이산화탄소 농도가 저하 되어 캘빈회로에서 루비스코가 옥시게나아제로 작용하여 이산화탄소 대신 산소를 RuBP와 결합시키므로 광합성에서 만들어지는 PGA가 반으로 줄어들어 광합성 효율이 낮아지며 이런 일련의 과정을 통하여 광 조건에서 산소를 소모하고 이산화탄소를 배출하는 광 호흡을 하게 된다. 광 호흡은 여름철 기온과 광도가 높고 건조할수록 증가하며, 일반적으로 한여름 광합성 효율이 낮아지는 이유는 기공이 닫혀 이산화탄소 흡수가 억제되고, 체내에 있는 산소를 소모하는 호흡을 하게 됨으로써 광합성 효율은 극히 낮아져 정상적인 생육을 하기가 어려워진다(Woo 등, 2014). 극고온과 광포화점 이상의 과다한 일사량을 효과적으로 적정 수준의 차광율로 차단함으로써 기공 폐쇄를 억제하여 기공이 열리도록 하고 기공전도도를 높임으로써 작물 성장이 가능한 환경을 조성해 줄 수 있다.
일반농가에서 여름철 온도 하강을 위해 보편적으로 사용하는 방법은 외부 기상환경 변화에 연계되지 않은 하루 종일 차광하는 전일 차광 방법을 이용하고 있다. 여름철에 흑색 차광망을 이용한 전일차광 방법은 작물의 생육 특성을 고려하지 않은 차광 방법으로 작물생육이 만족스럽지 못하다. 또한 전일차광은 차광율이 35∼40%로 여름철 시설 내 온도를 작물생육 온도까지 낮추기 어려우며, 장마철 또는 구름이 많을 경우에는 작물이 광 부족 현상으로 도장되거나, 낙과, 낙화가 되어 작물의 생산성을 저하시키는 요인이 되고 있다(Gent, 2007; Woo 등, 1995, 1996, 2014).
여름철 고온기 온실의 기온은 강제환기와 40∼50% 전일차광을 해도 시설과채류 생육최고한계 기온인 35℃ 초과한다. 이러한 전일차광의 단점을 보완한 것이 수시차광이다. 수시차광은 일사량이 필요 이상으로 높아지는 시기에 차광을 하여 여름철에 지나친 온도상승을 억제하여 고온기에도 작물의 개화, 수정불량, 광합성 저하 등 생육장해를 감소시켜 작물의 생산성을 향상시키고 고품질의 작물을 생산할 수 있도록 하는 방법이다. 수시차광에 이용되는 권장 차광율은 작물에 따라 다르나 일반적으로 50~70%이다. 차광율은 승온 억제 효과에 가장 크게 영향을 미치는 요인이다. 차광율이 커질수록 승온 억제 효과도 커진다. 효과적인 수시차광을 위해서는 작물별로 적정 차광율, 차광시기에 대한 세밀한 검토가 필요하다.
본 연구에서는 일반적인 50~70% 수시차광 보다 높은 90% 이상의 수시차광이 온실내의 작물 재배환경과 오이 엽온, Vcmax, Jmax, Thermal breakdown(high temp. inhibition), 광 호흡(Rd, day light leaf respiration)에 미치는 영향을 구명하고자 하였다.
재료 및 방법
본 연구는 전라북도 전주시 소재 한국농수산대학 플라스틱온실에서 2019년 6월 12일부터 8월 22일까지 71일간 수행하였다. 오이(장대명 백다다기기)를 5월 9일 파종하였고 6월 12일 코코피트 슬라브에 정식하였다. 첫 수확은 7월 17일이었다. 재배방법은 코코피트 수경재배로 하였으며, 배양액은 일본 야마자키 오이배양액 기준으로 조성하여 점적관수하였다. 차광은 정식 후 외부 일사량이 650W·m-2이상일 경우 자동으로 차광되도록 시스템(GIGA 스마트팜, KT)을 구성하였으며, 이와 같이 일사량에 따라 차광되는 방법을 수시차광이라고 하였다. 수시차광 온실은 전일차광에 이용되는 기존의 차광율 35∼40%보다 높은 차광 스크린 40%와 75% 보온스크린을 동시에 중복 차광(이후부터 90% 수시차광으로 표현)하여 90%이상의 차광효과를 내었다. 40% 차광 온실은 반자동으로 아침에 열고 저녁에 닫았다. 온실 측면은 광이 투과되도록 하였다. 측창은 개방하여 외부공기가 통풍되도록 조치하였다. 수시차광 시에는 천창과 측창을 열어 자연환기가 되도록 하였다. 실험온실은 2연동 PO필름 피복 온실 (90%이상 차광), 단동 PO필름 피복 온실 2동(40% 차광, 무차광)으로 하였다. 90% 차광과 40% 차광은 은색 알루미늄 증착 망을 이용하였다. 온실의 기온과 상대습도는 온실 중앙에서 지상 1.2m부근 3개소에서 측정한 평균값을 이용하였으며, 외부 일사량은 온실 동고 위에서 측정하였으며, 온실 내부 일사량은 중앙 1.2m 높이에서 측정하였다. 엽온은 생육이 가장 왕성한 엽령인 25∼30일 되는 6개체를 선정하여 측정하였으며, 엽온과 기온 간의 차를 분석하였다. 재배 기간 중 위치별 광량과 온도 측정은 data logger(CR3000, Campbell, Inc., USA)와 HOBO 소형 측정기(U23-001, Onset comp., USA)를 사용하였다.
Arrhenius function, Vcmax(Maximum carboxylation rate), Jmax(Maximum electron transport rate), thermal breakdown(High temperature inhibition), Rd(Day light leaf respiration) 값은 식 (1), (2), (3), (4), (5)의 모델식을 이용하여 계산하였으며, SAS(SAS Institute, Cary, North Carolina, USA)프로그램에서 제공하는 숫자함수, 산술연산자를 이용하여 작성한 계산 프로그램을 이용하였다.
Arrhenius function :
| $$f(T_L)=\exp\lbrack\frac{\triangle H_a}{298.15ℜ}(1-\frac{298.15}{T_L})\rbrack$$ | (1) |
여기서, TL : 엽온, Ha : 활성에너지[J·mol-1], ℜ : the universal gas constant[8.314JK-1·mol-1]
Rubisco maximum carboxylaton rate :
| $$V_{cmax}=\frac{V_{m,25}\;\exp\lbrack0.088(T_L-25)\rbrack}{1+\exp\lbrack0.29(T_L-41)\rbrack}$$ | (2) |
여기서, Vm, 25 : 25℃에서의 루비스코 카르복실화 속도(µmol·m-2·s-1)
Maximum electron transport rate :
| \begin{array}{l}J_\max=J_{\max25}\;f(T_L)f_H(T_L)\\J_{\max25}=1.67V_{cmax25}\end{array} | (3) |
여기서, fH : 비활성에너지[J·mol-1]
Thermal breakdown(High temperature inhibition) :
| $$f_H(T_L)=\frac{1+\exp(\frac{298.15\triangle S-\triangle H_d}{298.15\mathfrak R})}{1+\exp(\frac{\triangle ST_L-\triangle H_d}{\mathfrak RT_L})}$$ | (4) |
여기서, S : 엔트로피(JK-1·mol-1), Hd : 비활성에너지[J·mol-1]
Leaf respiration under day light :
| $$R_d=\frac{R_{d,25}\;\exp\lbrack0.069(T_L-25)\rbrack}{1+\exp\lbrack1.3(T_L-55)\rbrack}$$ | (5) |
여기서, Rd, 25 : 25℃에서의 잎의 호흡량(µmol·m-2·s-1),
TL : 엽온
통계분석을 위하여 시험구는 오이를 완전임의 배치법 3반복으로 배치하였으며, 통계분석은 SAS 프로그램을 이용하여 95% 신뢰수준에서 Duncan’s Multiple Range Test(DMRT) 검정을 실시하였다.
결과 및 고찰
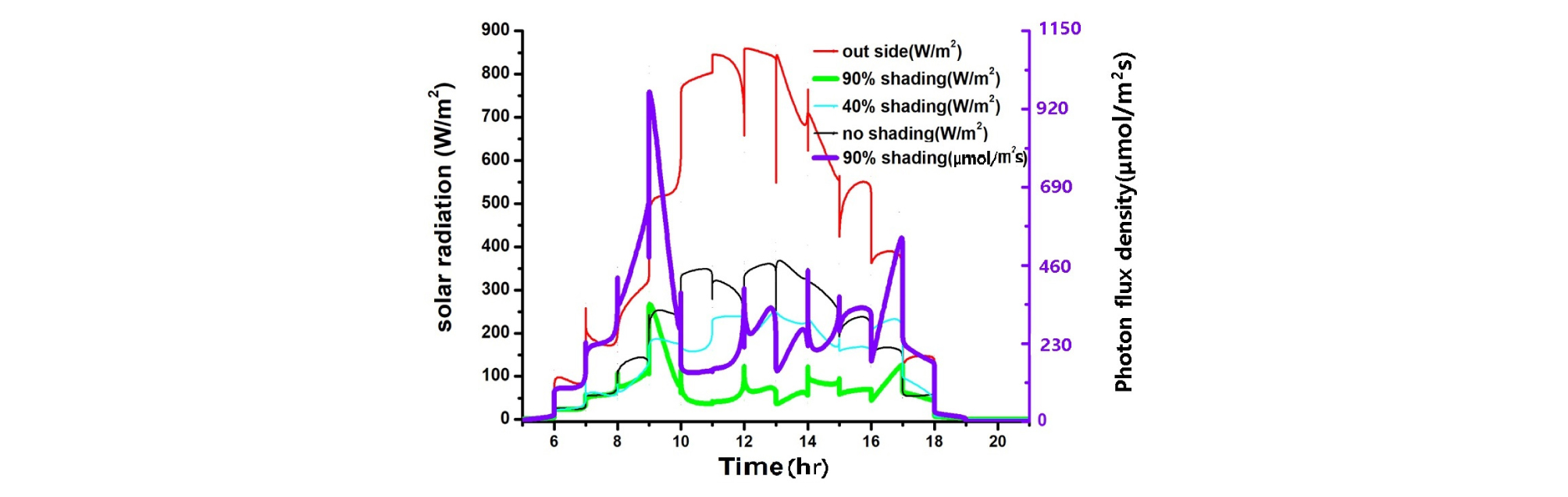
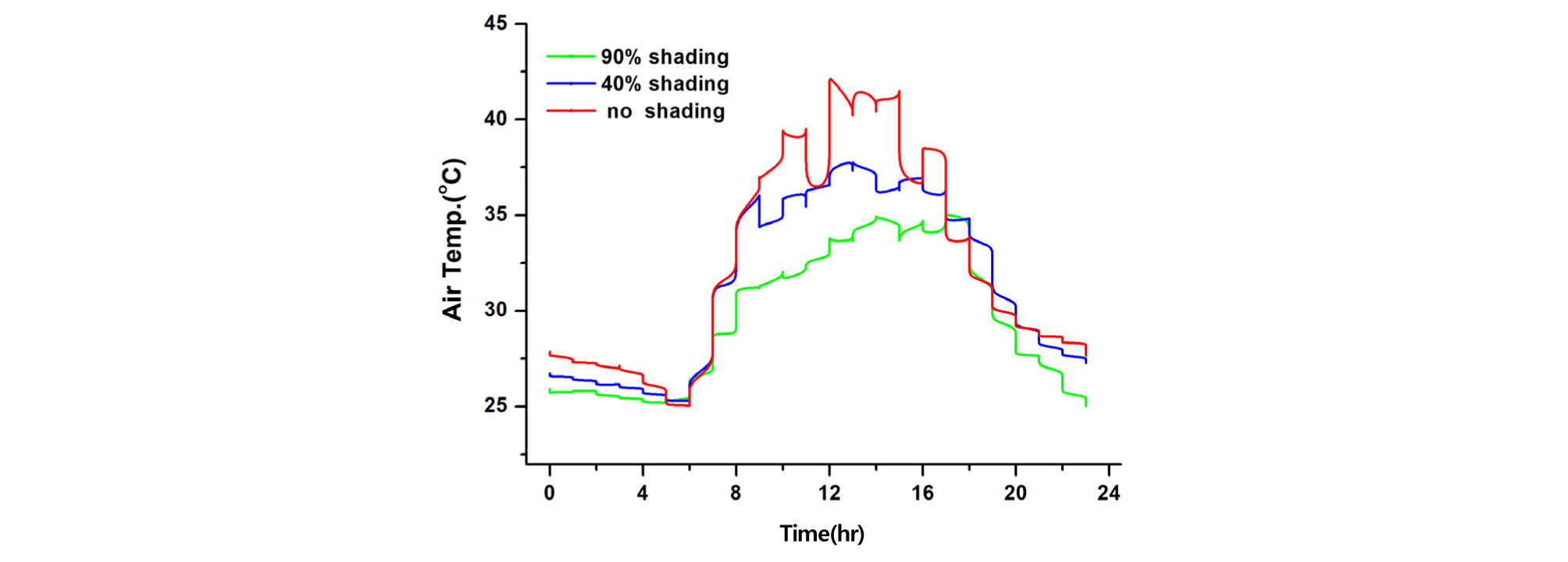
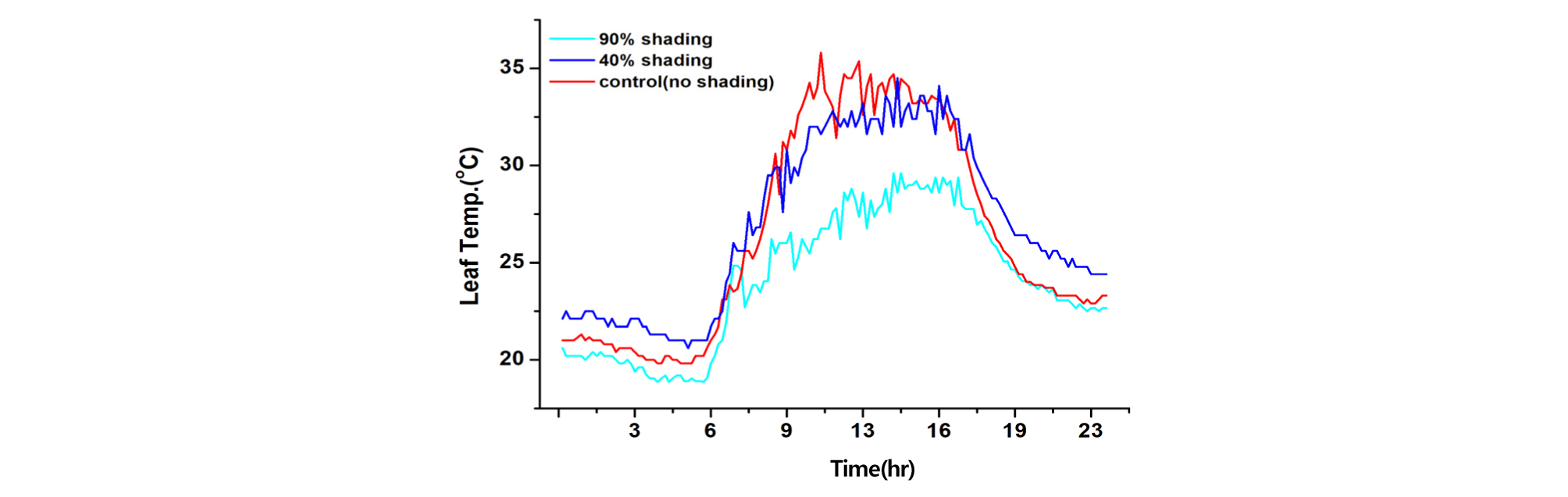
온실 내 환경을 측정한 결과, 90% 수시차광 온실이 40% 차광과 무차광 온실 보다 기온, 일사량, 엽온은 낮고, 엽기온차는 작은 반면, 상대습도는 높게 나타났다(Table 1, Fig. 1, 2, 3, 4). 기온은 90% 차광 온실이 33.6℃, 40% 차광 온실이 39.7℃, 무차광 온실이 41.2℃ 이었고, 엽온은 90% 차광 온실이 28.2℃, 40% 차광 온실이 32.6℃, 무차광 온실이 33.8℃로 차광율이 높을수록 기온과 엽온은 낮아졌다. 반면에 상대습도는 90% 온실이 40.4%, 40% 차광 온실이 32.9%, 무차광 온실이 28.9%로 차광율이 높을수록 높아졌다. 일사량은 90% 차광 온실이 68.2 W·m-2, 40% 차광 온실이 213.7 W·m-2, 무차광 온실이 280.5 W·m-2 순이었다. 광량은 오전 10시 전후와 오후 4시 이후 460 μmol·m-2·s-1이상을 보여, 90% 수시차광을 하여도 조도를 환산한 광량자속으로는 광보상점 23∼46 μmol·m-2·s-1 보다 상당히 높은 230∼460 μmol·m-2·s-1를 나타내었다. 이 값은 오이의 광포화점 1265 μmol·m-2·s-1보다는 낮으나 상추의 광포화점 632 μmol·m-2·s-1 보다 조금 낮은 수준의 광량임을 알 수 있었다(Fig. 1, Table 1).
Table 1.
Effect of different shading on air temperature, relative humidity, leaf temperature, solar radiation in greenhouse (Average value from 10 am to 16 pm on 9 August 2019).
최대 카르복실화 속도(Rubisco maximum carboxylation rate, Vcmax)는 90% 차광이 129.3 μmol・m-2・s-1으로 다른 처리에 비해 낮았다. 최대 전자전달 속도(Maximum electron transport rate, Jmax)는 90% 차광과 무차광이 각각 184.6 μmol・m-2・s-1, 183.9 μmol・m-2・s-1로 비슷한 값을 나타내었고 40% 차광이 190 μmol・m-2・s-1으로 높은 값을 나타내었다(Table 2). Jmax(최대 전자전달 속도) 값이 100 μmol・m-2・s-1이하여야 하나 비정상적인 조건으로 작물 체내 원활한 대사과정이 일어나지 않고 있음을 의미하며(Collatz 등, 1991; Farquhar 등, 1980; Huitzimengari 등, 2014; Jackson 등, 1981), 오히려 90% 차광이 Vcmax가 100에 가까운 양호한 값을 보임을 알 수 있다(Collatz 등, 1991; Farquhar 등, 1980). Jmax 모델식은 주로 기온이 25℃를 기준으로 하고 있으므로 극고온에 대한 모델식은 오차가 예상되므로 고온에 대한 새로운 모델식이 필요함을 나타내고 있다.
Table 2.
Effect of different shading on Rubisco maximum carboxylation rate (Vcmax), maximum electron transport rate (Jmax), Arrhenius function (f(TL)), thermal breakdown (fH(TL)) and leaf respiration under day light (Rd) by mobile shading (Average value from 10 am to 16 pm on 9 August 2019).
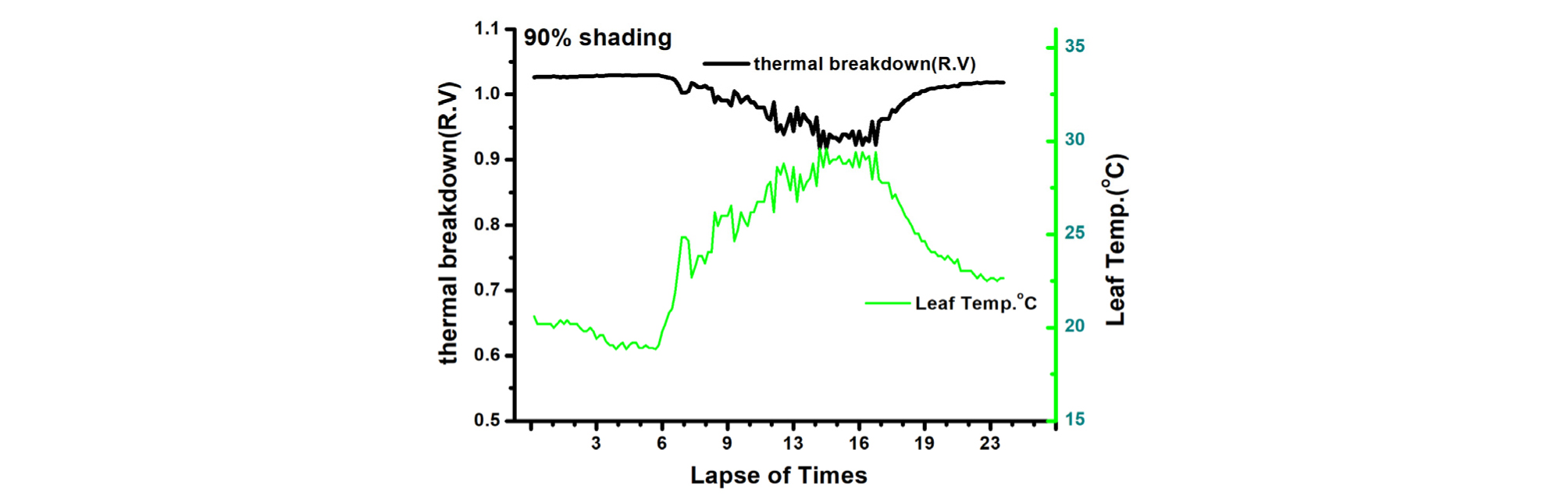
Thermal breakdown(high temp. inhibition)은 무차광이 0.724(relative value; R.V), 40% 차광이 0.793(R.V.), 90% 수시차광이 0.951(R.V.)로 나타나 90% 수시차광이 유의하게 높은 수준의 thermal breakdown값을 보여 그 이하 값에서는 정상적인 생육 재배가 가능하다는 의미이므로 90% 수시차광 시 고온에 의한 생육 억제 현상은 본 실험조건 하에서 가장 적을 것으로 판단되었다. 광 호흡을 보면 무차광이 2.754 μmol・CO2・m-2s-1, 40% 차광이 2.534 μmol・CO2・m-2s-1, 90% 수시차광이 1.870 μmol・CO2・m-2s-1으로 수시차광이 타 처리에 비해 유의하게 낮았다. 그동안 90% 수시차광은 효과적으로 광 호흡을 억제할 것이라는 추측(Woo 등, 2014)이 객관적인 수치로 나타났다. 본 실험조건 아래에서는 여름철 고온기에 90% 이상의 수시차광이 타 처리보다 확실히 광호흡 억제와 더불어 고온장해 없이 오이 작물 재배에 양호한 환경조건을 조성하는 것으로 밝혀졌다(Table 2).
Arrhenius function f(TL) 값은 무 차광> 40% 차광> 90% 차광 순이었다(Table 2). Arrhenius function 값은 온도에 따른 생육 반응 정도를 나타내는 것으로 그 값이 클수록 온도에 따른 생육 반응이 예민하게 반응한다는 의미로 90% 차광이 타 처리에 비하여 온도에 둔감하게 생육 반응이 일어남을 나타내고 있다.
엽온(leaf temperature)과 Thermal breakdown관계를 보면, 90% 차광조건에서 Thermal breakdown 값은 0.9 이상이고 엽온은 30℃ 이하가 되어 고온에 의한 생육억제 현상이 적음을 알 수 있으며(Table 2, Fig. 4), 다른 처리에서는 정도의 차이는 있으나 Thermal breakdown 값이 0.79, 0.72로 모두 고온에 의한 생육이 억제됨을 나타내고 있다(Table 2, Fig. 5, 6).
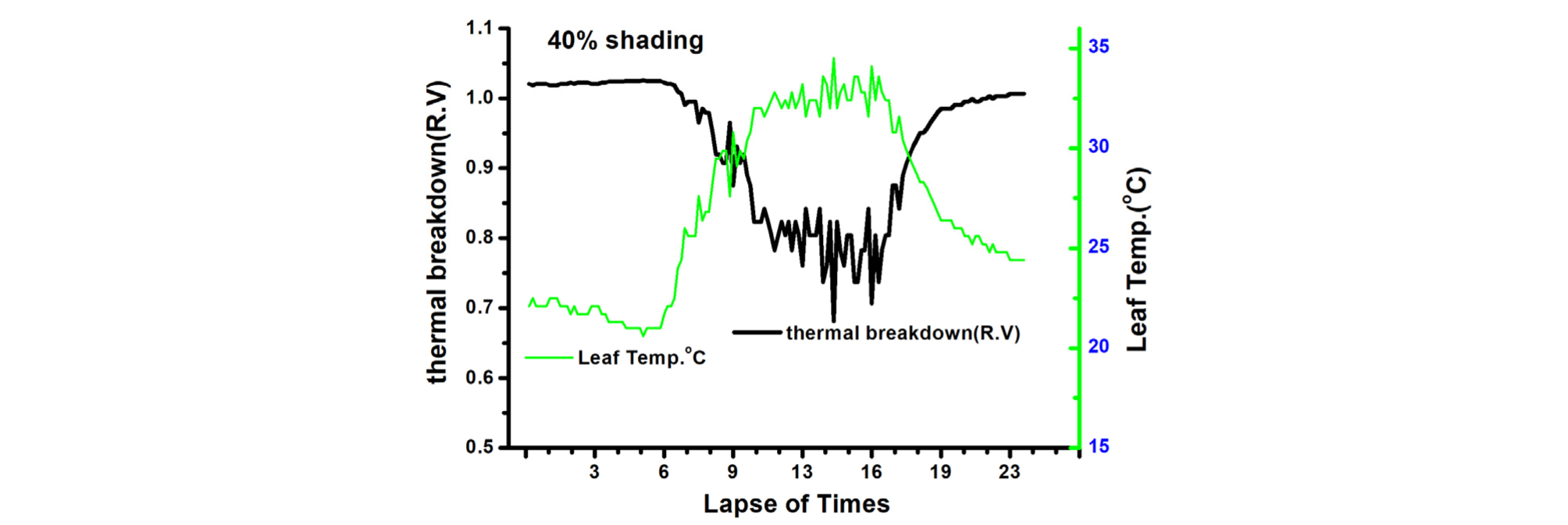
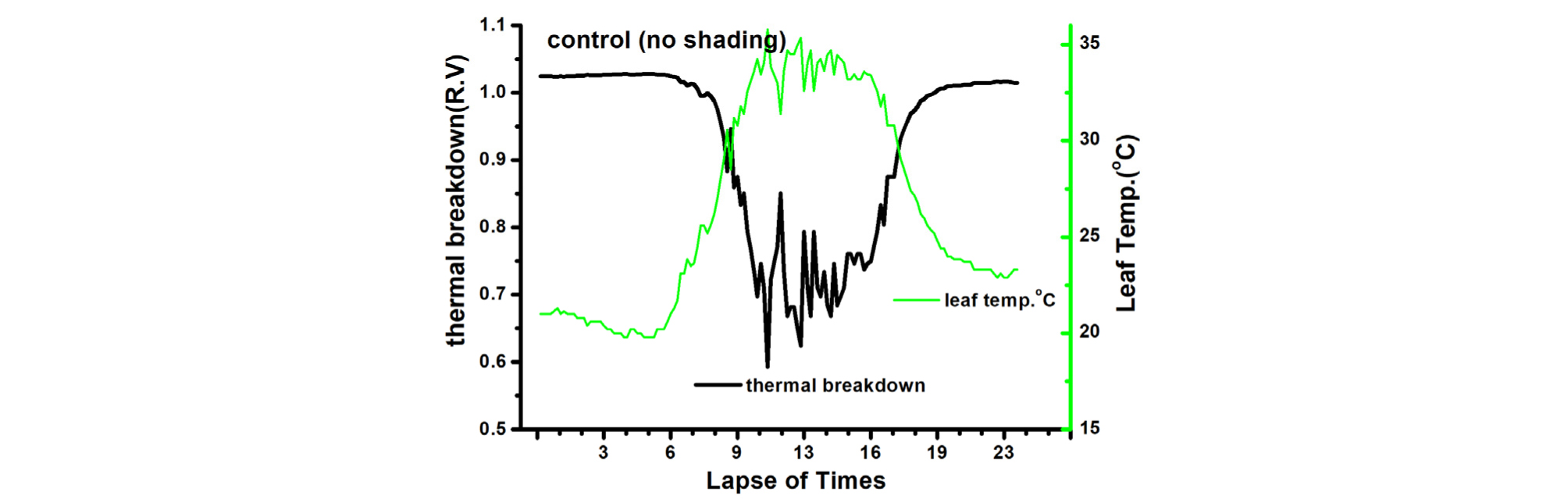
엽기온차는 엽온과 기온 간의 차이에 따른 작물의 스트레스를 나타내는 지표(Crop Water Stress Index; CWSI)로 이용되고 있다. 엽기온차와 Thermal breakdown 관계를 보면 엽온-기온차이가 +(plus)상태이면 Stress 상태이고 엽온-기온 차이가 -(minus)상태이면 non-stress 상태(Idso 1982; Jackson 등, 1981, Woo 등, 2000)를 표시한다(Fig. 7, 8, 9, 10). 일반적으로 기공전도도가 높고 증산작용이 활발하게 일어난다는 것은 기공이 열린 상태에서 작물 체내에 있는 수증기가 외부 배출되므로 엽온이 낮아진다는 것을 의미한다(Idso 1982; Jackson 등, 1981; Woo 등, 2000). 다시 말하면 엽기온차는 엽온(leaf temperature)에서 대기 온도(air temperature)를 뺀 값으로 일반적으로 증산작용이 원활하게 일어나면 엽온은 낮아지게 되어 엽온-기온의 값은 마이너스(-)가 된다. 엽온-기온의 값이 마이너스(-)가 되면 그 작물은 Non stress 상태가 된다는 것을 의미한다. 엽온-기온의 값이 플러스(+)가 되면 기공전도도가 낮아져 증산작용이 억제되어 엽온이 기온보다 높아진 상태이며 이런 상태를 Stress를 받았다고 할 수 있다(Idso 1982; Jackson 등, 1981). 이처럼 엽온-기온의 값이 플러스(+)가 되면 기공 닫힌 상태이거나 기공전도도가 감소된 상태임을 추정할 수 있다. 따라서 마이너스(-)값이 클수록 오이의 수분 스트레스 정도는 낮다고 할 수 있으며, 플러스(+) 값이 클수록 오이 수분 스트레스는 높다고 할 수 있다.
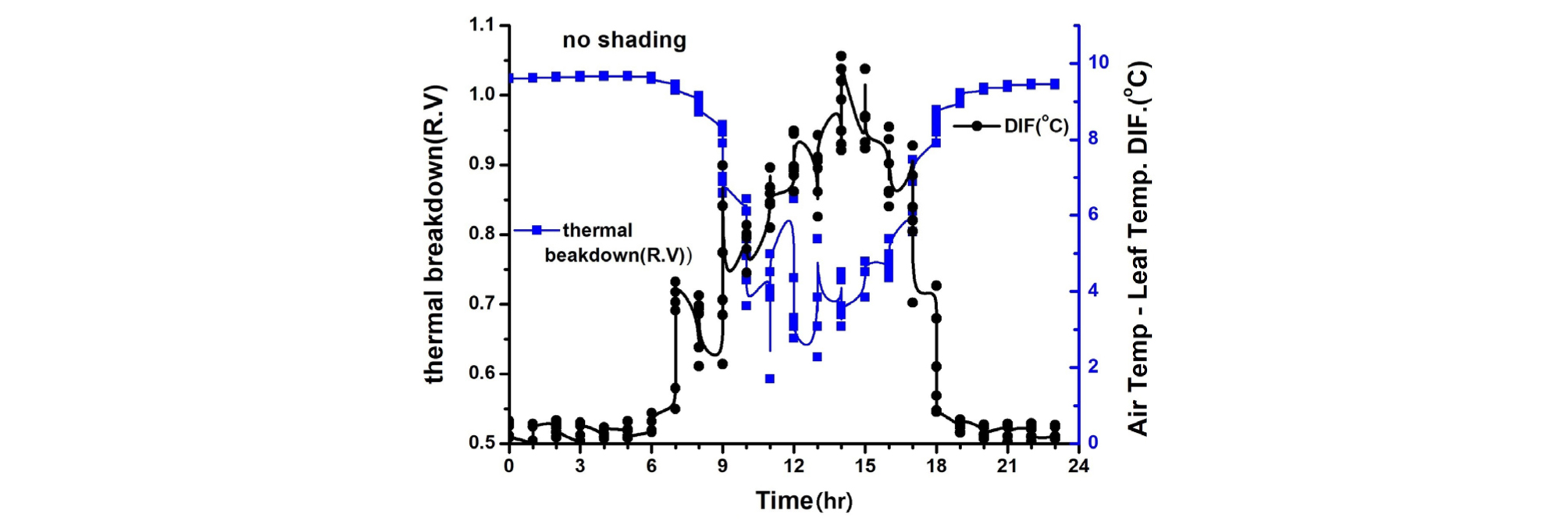
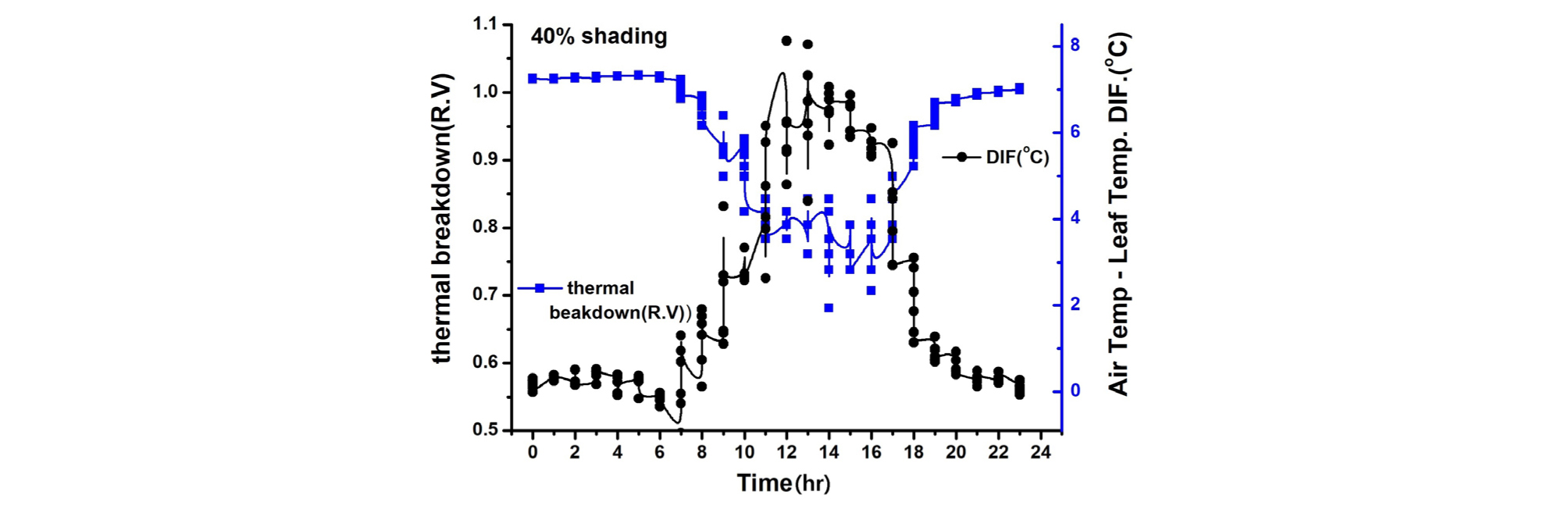
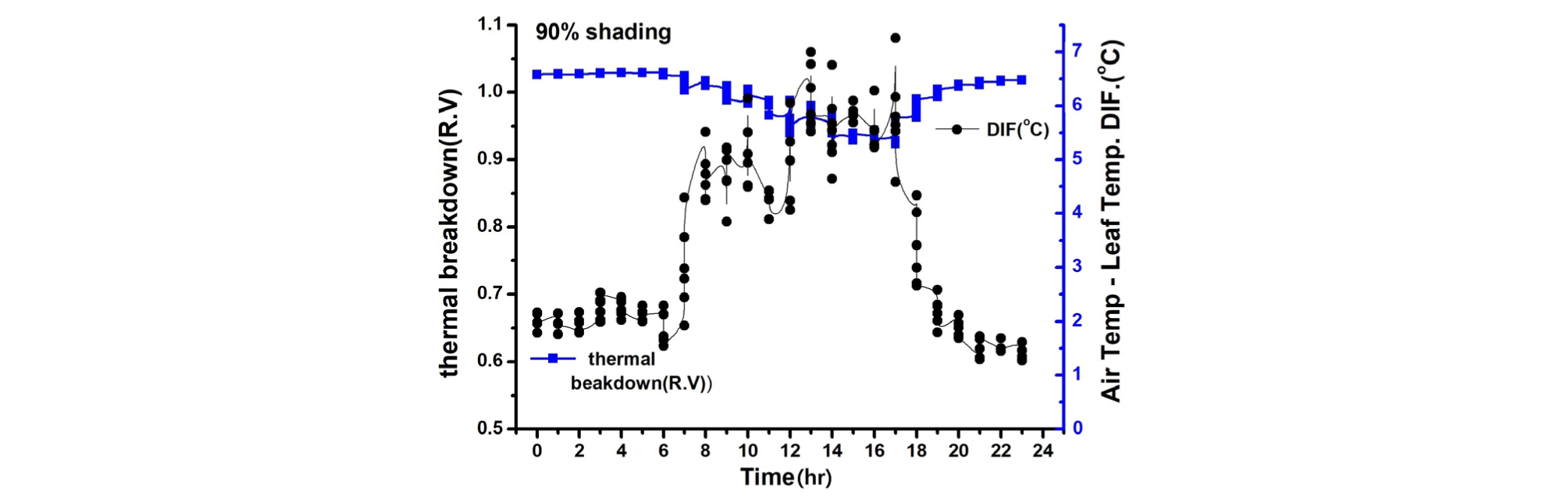
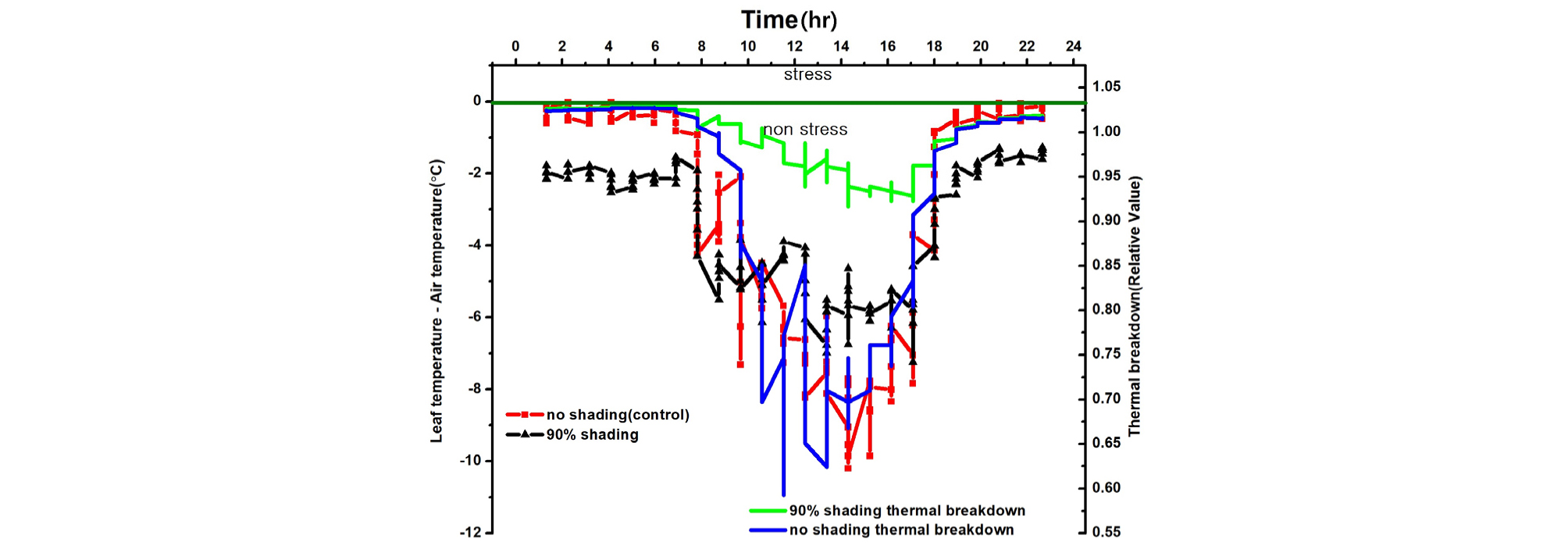
본 연구에서 엽기온의 차의 관계에서 엽온이 기온보다 낮은 -방향에서는 Non stress이며, 엽온이 기온보다 높은 +상태에서는 Stress 상태로 설정한 Idso(1982)과 Jackson 등(1981)의 연구결과를 근거로 분석해보면 90% 수시차광이 다른 처리에 비해 엽기온차가 –5.4℃. 40% 차광이 –7.1℃, 무차광이 –7.4℃로 기존의 연구(Woo 등, 2014)와는 다른 결과가 나타났다(Fig. 7). 이는 차광율이 높을수록 엽기온차가 크게 난 기존연구 결과와 다르나 이는 실내 기온이 극고온 상태이고 근권부 토양수분 차이에 기인하는 것으로 추측되나 더욱 연구할 과제라 생각된다. 본 연구결과를 기준으로 Thermal breakdown과 연계하여 분석해보면 40% 차광과 무차광조건에서는 극고온기 엽기온차가 Thermal breakdown 값 보다 높아 고온장해를 받는 것으로 나타났다(Fig. 7, Fig. 8). 그렇지만, 90% 차광에서는 대부분 엽기온차 값이 극히 일부분을 제외하고는 Thermal breakdown 값보다 작아 고온장해를 받지 않는 것으로 나타났다(Fig. 9, Fig. 10). 따라서 엽기온차로 보면 Non stress 상태라 할지라도 Thermal breakdown 값을 기준으로 보면 장해를 받는 것으로 나타났으며, 10℃ 이상의 엽기온차가 장시간 지속적으로 반복하여 일어날 때 잎에서의 결로로 인해 병해 발생이 증가할 우려가 있다. 이에 대해서는 좀 더 깊이 있는 연구와 분석이 필요하다고 생각된다. 또한, 온실의 지나친 온도상승을 억제하여 고온기에도 작물의 개화, 수정불량, 광합성 저하 등 생육장해를 방지하여 작물의 생산성을 향상시키고 고품질의 작물을 생산할 수 있도록 90% 차광율로 수시차광을 실시할 경우 차광 커튼이 닫히는 일사 기준에 대한 세밀한 연구가 뒷받침되어야 할 것으로 판단된다.
