

서 론
온도는 식물의 생육과 발육에 영향을 주는 중요한 환 경요인으로, 모든 생물학적 과정은 온도에 반응하며, 최 저, 최적 및 최고 온도로 나뉜다(Jami Al-Ahmadi과 Kafi, 2007; Porter와 Gawith, 1999; Tokumasu 등, 1985; Yan 과 Hunt, 1999). 최저 온도는 작물의 재배가 시작되는 시기를 결정하며, 최고 온도는 고온기의 재배적 영향을 고려하는데 지표가 된다. 주요 온도(최저, 최적 및 최고 온도)로 식물체의 발육 현상을 예측할 수 있는데, 주로 재배 환경의 변화가 심하지 않은 발아실험을 통해 예측 할 수 있다(Roché 등, 1997). 보통, 발아율은 최저와 최 적온도 사이에는 증가하다가 최적과 최고 온도 사이에는 감소한다. 그리고 최저 이하와 최고 온도 이상에서는 발 아율이 정지된다. 주요 온도는 최저 온도 이하나 최고 온 도 이상의 환경을 추정하는데 중요하다(Jami Al-Ahmadi 과 Kafi, 2007; Montieth, 1981; Seefeldt 등, 2002). 따 라서, 주요 온도는 이상기후 환경에 대한 작물의 반응을 예측하는데 중요한 정보를 제공해 줄 수 있다. 또한 작 물의 생육과 발육을 위한 주요 온도는 재배, 생육과 성 숙 및 수량을 예측하는데 중요한 정보로 활용될 수 있다.
주요 온도를 예측하는데 모델식을 이용하여 예측하려 는 많은 연구들이 진행되어 왔다(Aflakpui 등, 1997; Brown과 Mayer, 1988; Carberry와 Abrecht, 1990; Del Monte와 Tarquis, 1997; Hardegree, 2006; Iannucci 등, 2000; Jami Al-Ahmadi과 Kafi, 2007; Roché 등, 1997; Seefeldt 등, 2002). 선형모델을 이용한 연구(Aflakpui 등, 1997)와 쌍곡선 모델을 이용한 연구(Dumur 등, 1990; Hardegree, 2006; Roché 등, 1997; Seefeldt 등, 2002; Yan과 Hunt, 1999; Yin 등, 1995) 및 비선형함수인 베 타함수(beta distribution) 모델을 이용한 연구들이 있었다 (Yan과 Hunt, 1999; Yin 등, 1995). 특히, 베타함수는 작 물의 온도에 따른 효소 반응처럼 작물 발육량에 따른 온도를 함수로 표현한 것이다(Yan과 Hunt, 1999). 이 함 수는 생물학적 과정에 적용될 수 있는 함수로 알려져 있다(Yan과 Hunt, 1999; Yin 등, 1995).
대부분 작물의 최저 온도와 최적 온도는 각각 0~5°C 와 20~25°C 범위이다(Monteith, 1981). 상추는 호냉성 작물로, 발아한계 최저 온도는 4°C이고, 최적 온도는 15~20°C, 최고 온도는 25°C이다. 작물 생육에 필요한 주 요 온도는 품종별 생육단계별로 요구되며(Del Monte와 Tarquis, 1997; Iannucci 등, 2000; Jami Al-Ahmadi과 Kafi, 2007; Monteith, 1981; Olivier와 Annandale, 1998; Seefeldt 등, 2002; Slafer와 Savin, 1991; Tan 등, 2000), 발아 실험을 통해 주요 온도를 예측할 수 있다.
이 연구의 목적은 상추 발아 실험을 통해 주요 온도 (최저, 최적, 최고)를 선형, 쌍곡선 및 베타 함수 등식을 이용하여 예측하고 3가지 함수를 비교하는 것이다.
재료 및 방법
실험 재료는 상추 품종 중 치마상추인 흑상추(Aram Seed. Co.)을 이용하였다. 발아상은 항온항습이 되는 생 육상을 이용하였다. 온도 설정은 10, 14, 16, 20, 24, 28, 32와 36°C로 항온조건에서 발아시켰다. 발아실험은 9cm 페트리디쉬에 No. 2 필터페이퍼를 2장 깔고, 그 위 에 100개의 종자를 올려놓았다. 증류수 10mL를 넣고 항온항습실에서 발아를 시켰다. 유근이 1mm 나왔을 때 를 발아로 정하였으며, 매일 발아한 종자를 제거하였다.
발아는 로지스틱 함수(등식 1)를 이용하여 계수를 찾 았다.
| $$\mathrm Y\;=\;\mathrm M\;\times\;{\lbrack1\;+\;\exp(-\mathrm k\;\times\;(\mathrm t\;-\;\mathrm L))\rbrack}^{-1}$$ | (1) |
여기에서, Y는 t시간에 총발아한 개수, M는 잠재적인 발아수, L는 시간 상수, 그리고 k는 증가율(Roché 등, 1997)이다. 주요 온도는 온도에 따른 발아율이 50%된 시점의 역수(1/GR50)로 예측하였다. 주요온도를 예측하기 위해 선형함수, 쌍곡선 함수와 베타함수를 이용하였다 (Cho et al., 2009). 함수의 계수들은 Gauss-Newton 알 고리즘을 이용하여 예측하였다. 통계분석은 SigmaPlot (SPSS Inc., Korea)와 SAS(Statistical Analysis System) 프로그램을 이용하였으며, 완전임의배치법으로 4반복 실 시하였다. 실험결과들은 ANOVA(analysis of variance) 검정하였다.
결과 및 고찰
항온 상태하에서 처리한 상추의 종자 발아 곡선은
Fig. 1과 같다. 온도가 20°C와 24°C 처리구에서 발아속 도가 빨랐으며, 32°C 처리구가 빨아속도가 가장 느렸다. 온도 처리별 발아시는 10°C와 32°C 처리구가 3일, 14°C 처리구가 2일, 16°C, 20°C, 24°C, 28°C 처리구가 1일이 었다. 28°C와 32°C 처리구를 제외하고 다른 처리구에서 높은 발아율을 보였다.
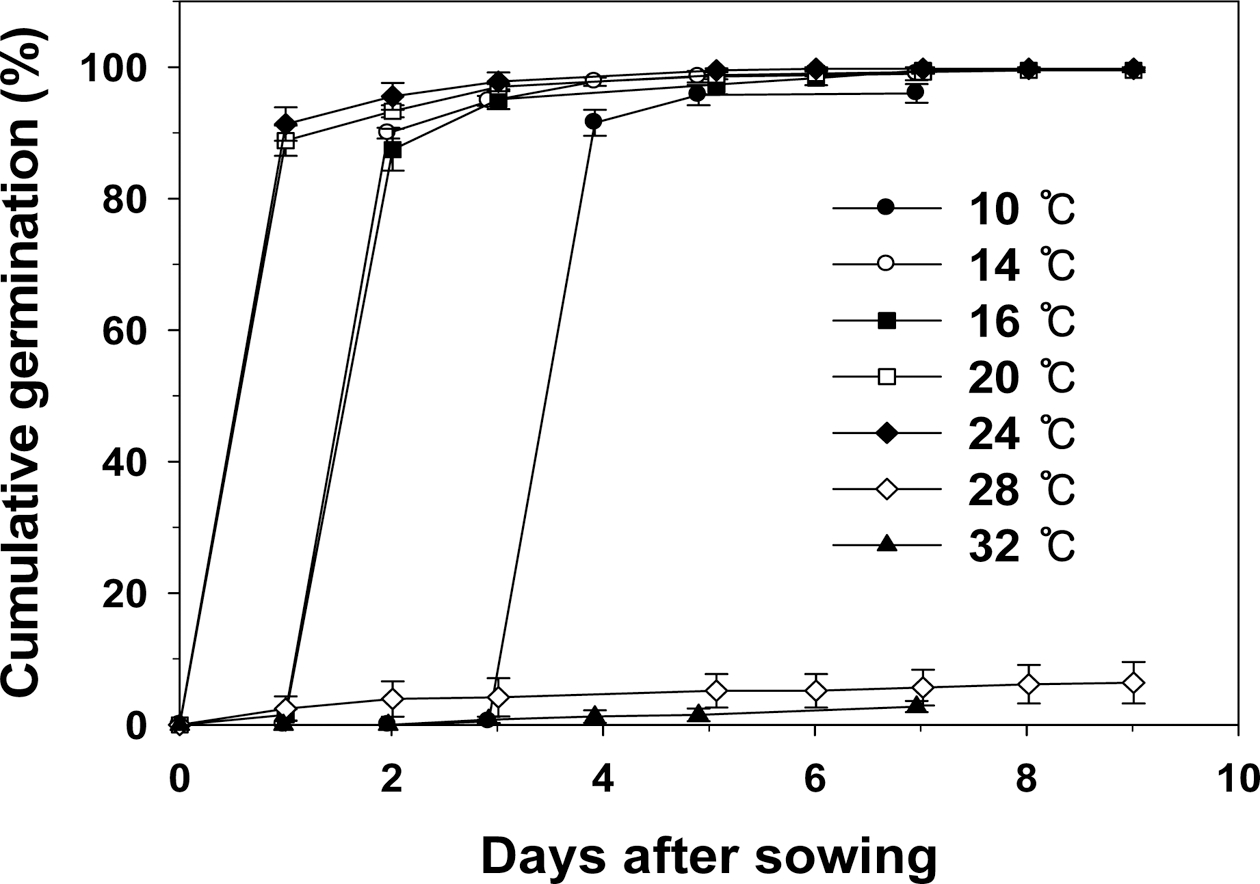
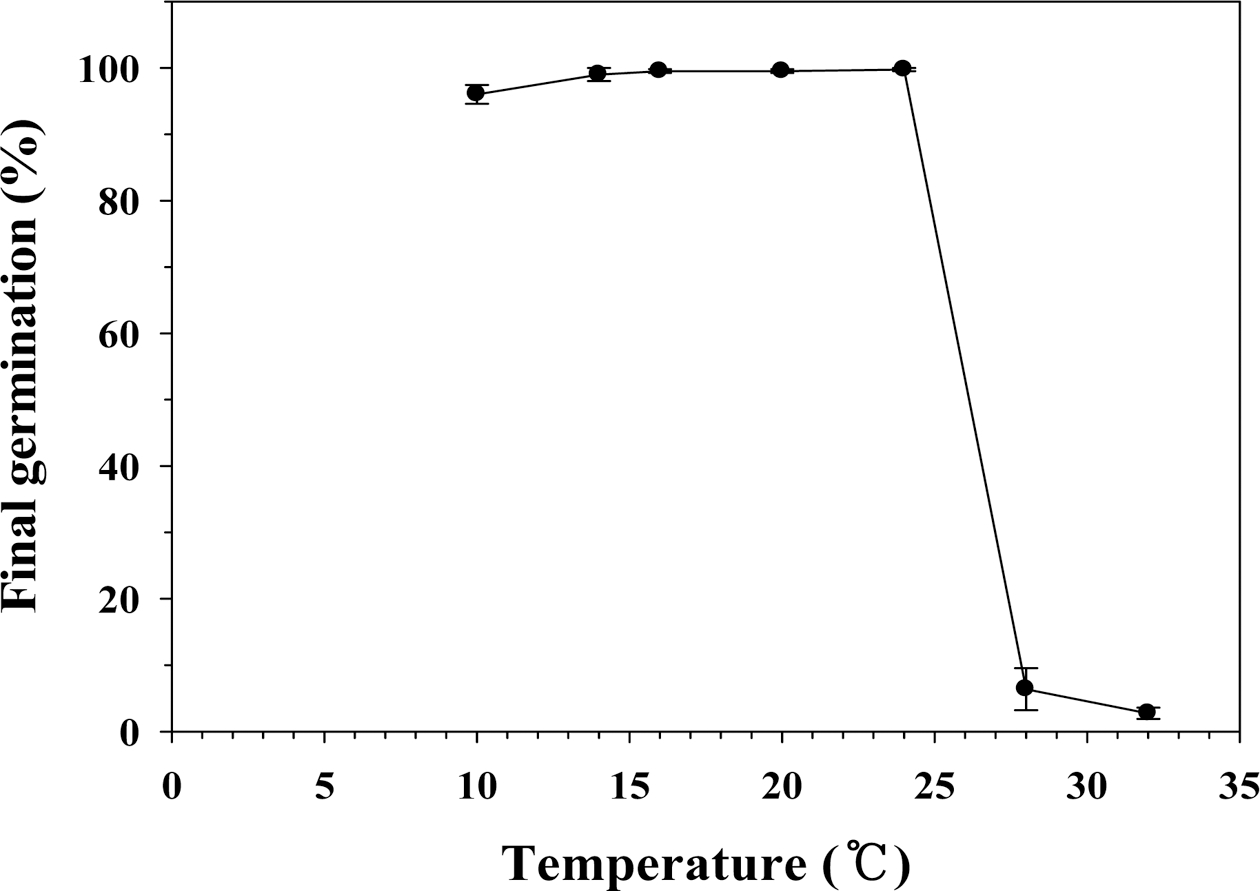
최종 발아율은 28°C와 32°C 처리구를 제외하고 다른 처리구에서 높은 최종 발아율을 보였다(Fig. 2). 10°C, 14°C, 16°C, 20°C, 24°C 처리구간 최종 발아율의 차이는 없었다. 따라서 10°C에서 24°C까지의 최종 발아율에는 문제는 없으나, 발아시와 발아속도를 고려해 볼 때, 20°C 와 24°C가 상추가 발아하는데 적당한 온도라는 것을 알 았다. 상추는 호냉성 채소로 알려져 있다. 일반적으로 호냉성 채소의 발아온도는 12.8~18.3°C로 알려져 있다. 상추의 발아한계 최저 온도는 4°C이고, 최적 온도는 15~20°C, 최고 온도는 25°C로, 본 연구에서의 최저, 최 적, 최고온도 범위는 각각 3.7~7.9°C, 19.5~23.3°C, 28.0~ 32.0°C이었다. 상추의 주요 온도는 품종별로 다를 것으 로 판단되며, 각 품종별 주요 온도를 찾는 것이 재배관 리적인 측면에서 중요한 정보를 제공해 줄 것이다.
항온 처리에 따른 발아모델의 계수값들은 Table 1과 같다. 10°C에서 24°C 처리구간을 보면, 최대발아율(M) 과 증가율(k)간에는 차이가 없었으나, 시간상수(L)는 10°C 처리구에서 가장 높은 수치를 보였으며, 20°C와 24°C 처리구에서 가장 낮은 수치를 보였다.
Table 1.
Coefficients of the germination model on different constant temperatures.
| Temp. (°C) | Logistic coefficientz | Pr > F | ||
|---|---|---|---|---|
| M | k | L | ||
| 10 | 95.9 ± 0.70 | 8.3 ± 2.02 | 3.6 ± 0.09 | <0.0001 |
| 14 | 97.5±0.46 | 10.7±3.32 | 1.7±0.72 | <0.0001 |
| 16 | 98.2±0.59 | 6.2±0.94 | 1.7±0.05 | <0.0001 |
| 20 | 98.0±0.48 | 9.9±2.64 | 0.8±0.61 | <0.0001 |
| 24 | 98.8±0.49 | 11.0±6.35 | 0.8±1.32 | <0.0001 |
| 28 | - | - | - | - |
| 32 | - | - | - | - |
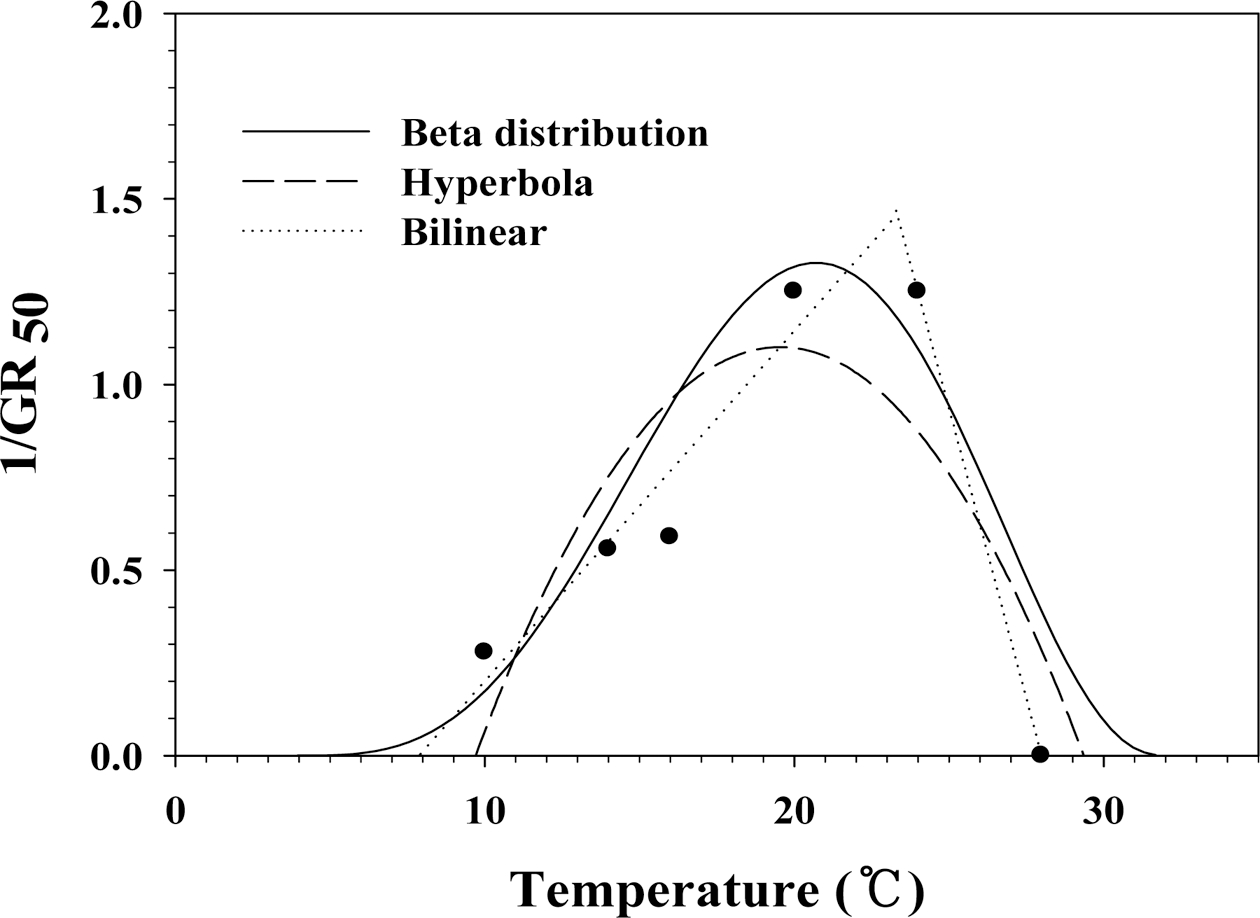
각 온도 처리별 50% 발아가 된 시점의 역수를 그래프 로 보면, Fig. 3과 같다. 28°C와 32°C 처리구는 50% 발 아율을 보이지 않아 28°C 처리구를 0값으로 하여 계산 하였다. 등식 2a, 2b, 3, 4a를 이용하여 주요 온도를 조 사하였다(Table 2). 3가지 함수를 이용한 경우, 평균 최 저, 평균 최적, 평균 최고 온도 및 평균 온도범위차는 각각 7.1°C, 21.2°C, 29.8°C, 22.7°C였다. 선형 함수(등식 2a와 2b)를 이용할 경우, 최저, 최적, 최고온도 및 온도 범위차는 각각 7.9°C, 23.3°C, 28.0°C, 20.1°C였으며, 쌍 곡선 함수(등식 3)를 이용할 경우, 최저, 최적, 최고 온 도 및 온도범위차는 각각 9.7°C, 19.5°C, 29.4°C, 19.7°C 였으며, 베타 함수(등식 4a)를 이용할 경우, 최저, 최적, 최고 온도 및 온도범위차는 각각 3.7°C, 20.7°C, 32.0°C, 28.3°C였다.
Table 2.
The cardinal temperature for 50% germination of spinach seed using linear, parabolic, and beta distribution equations.
| Equation | Temperaturez (°C) | |||
|---|---|---|---|---|
| Tmin | Topt | Tmax | Difference | |
| Linear | 7.9 | 23.3 | 28.0 | 20.1 |
| Parabolic | 9.7 | 19.5 | 29.4 | 19.7 |
| Beta distribution | 3.7 | 20.7 | 32.0 | 28.3 |
| Mean | 7.1 | 21.2 | 29.8 | 22.7 |
온도는 식물의 생장과 발육 및 수량을 예측하는 모델 에 사용되는 환경적인 변수 중 하나이다. 온도에는 주요 온도(최저, 최적, 최고 온도)가 있는데, 주요 온도는 다 양한 식물의 생물 계절학, 적응 및 수량을 계산하는데 중요하다(Yan과 Hunt, 1999). 또한 주요 온도는 유효적 산온도(growing degree-days, growing degree unit)를 계 산하는데 이용된다(McMaster와 Wilhelm, 1997). 이러한 유효적산온도는 생육과 수량을 예측하는데 이용되고(Tei 등, 1996), 시간(년도 또는 날짜)에 따른 생육과 수량 예 측 모델 함수를 개량하는데 이용될 수 있다(McMaster와 Wilhelm, 1997). 따라서, 주요 온도는 기후변화예측에 중 요한 자료로 활용될 수 있을 것이다.
발아모델에 있어서 선형 함수는 최적 온도를 기준으로 2개의 함수를 얻을 수 있는데, Craufured 등(1998)의 보 고에 따르면, 어떤 식물은 최고 온도가 198°C로 예측되 었다라고 하였다. 쌍곡선 함수는 최적 온도를 중심으로 좌우대칭으로 최저 온도 근처에서 불안정한 곡선을 보이 지 않는다(Yin 등, 1995). 최근 들어 온도에 따른 식물 체의 반응에 대해 베타 함수로 표기하는 연구가 진행되 고 있다(Yin 등, 1995). 이전의 등식과 비교하여 베타 함수는 몇몇 반응에 대해 실제적인 곡선을 나타내 주는 장점이 있다. 많은 연구들이 이 함수를 이용하고 있다 (Yan과 Hunt, 1999; Yin과 Kropf, 1996; Yin 등, 1995, 1996). Yan과 Hunt(1999)는 생식질(germplasm) 조사나 검사, 작물 생육 모델링, 인공광 이용 작물 생산 시스템 에 적용될 수 있다고 하였다.
