

서 론
수박 접목묘 생산 기술은 병충해 저항성을 강화하고 토양 전염성 병을 효과적으로 제어할 수 있어 중요한 기술로 자리 잡고 있다(Bie 등, 2017; Maurya 등, 2019; Bantis 등, 2020; Kwack 등, 2021). 수박 접목묘는 연작 장해를 극복할 뿐만 아니라 생산량과 품질을 높이고, 비생물적·생물적 스트레스에 대한 저항성을 향상시키는 데 기여한다(Colla 등, 2010; Schwarz 등, 2010; Ertle와 Kubota, 2022). 재배와 육묘의 분업화가 가속화되면서 전문적인 공정육묘 시스템의 수요와 규모는 지속적으로 확대되고 있다(Kang 등, 2010; Park과 Runkle, 2017). 그러나 최근 이상기후로 인한 여름철 장기적인 장마로 일조량이 부족해지면서 규격화된 고품질 묘를 안정적으로 생산하기 어려운 상황이 발생하고 있다(Bisbis 등, 2019). 이러한 문제를 해결하기 위해 연중 안정적으로 고품질 규격묘를 생산할 수 있는 식물공장형 육묘시스템 기술이 주목받고 있다(Jang 등, 2014).
식물공장형 육묘시스템은 광도, 온도, 습도, CO2 등 다양한 환경 요인들을 정밀하게 제어할 수 있으며, 외부 환경의 영향을 받지 않기 때문에 연중 균일한 고품질 규격묘를 관행보다 더 빠르게 생산할 수 있다는 장점이 있다(Park 등, 2020). 또한, 높은 자원 이용효율과 토지 면적대비 연간 생산성 증가로 인해 고품질 작물을 생산할 수 있는 가능성을 높이고 있다(Yang 등, 2018; Zheng 등, 2023). 현재 국내 대다수의 공정육묘는 자연광을 이용하는 유리온실 또는 플라스틱 온실에서 이루어지고 있으나, 이러한 온실은 광, CO2, 온도, 습도 등과 같은 외부 환경에 노출되어 매년 일정한 품질의 묘를 생산하기 어렵다(Um 등, 2009). 반밀폐 온실에서는 온실 구조와 식물 및 잎의 위치, 잎의 위치에 따라 수광량이 달라지므로, 식물의 광합성 효율이 일정하지 않다는 연구 결과가 보고된 바 있다(Wang 등, 2021). 게다가 기후변동성으로 인한 풍수해, 고온해, 냉해 등의 기상 재해가 증가함에 따라 고품질 규격묘 생산이 더욱 어려워질 것으로 예상된다.
빛 에너지는 식물이 광합성을 하는데 가장 중요한 요소 중 하나이다(Bednarczyk 등, 2020). 광도는 광합성 효율을 결정하는 주요 요인이므로, 식물이 필요로 하는 광을 적절히 공급하는 것이 매우 중요하다(Ghorbanzadeh 등, 2020). 식물 유묘의 경우, 낮은 광도는 효율적인 광합성을 제공하지 못해 식물이 도장하게 되고, 반대로 과도한 광도는 광호흡을 유도하여 광합성 효율을 저하시킨다(Ruangrak과 Khummueng, 2019). 따라서 식물의 생산성과 품질을 유지하기 위해 최적의 광도와 광질을 공급하는 것이 필요하다(Davis와 Burns, 2016; Shafiq 등, 2021).
LED가 식물 재배용 인공 광원으로 효과적이라는 연구 결과가 활발히 이루어지고 있으며, 이러한 연구는 엽채류 및 과채류의 생육과 품질에 LED가 미치는 영향을 폭넓게 분석하고 있다(Chen 등, 2022; Chen 등, 2023; Yun 등, 2023; Zheng 등, 2023). 이에 본 연구는 식물공장형 육묘시스템에서 광도가 수박 접목묘의 활착 이후 생육과 묘의 소질에 미치는 영향을 구명하기 위해 수행되었다.
재료 및 방법
1. 실험재료 및 식물공장 육묘시스템 내 환경
본 연구에서는 고온 육묘 시 널리 이용되는 접목묘 품종으로 대목은 불로장생(‘BullojangSaeng, BS’, Syngenta Korea Co., Ltd., Korea)을, 접수는 부라보꿀(‘BravoKkul, BK’, Nongwoobio Co., Ltd., Korea)을 선택하였다. 또한, 저온 육묘 시 널리 이용되는 품종으로 대목은 동장군(‘DongjangGun, DG’, Syngenta Korea Co., Ltd., Korea)을, 접수는 산타꿀(‘SantaKkul, SK’, Nongwoobio Co., Ltd., Korea)을 선택하였다. 대목은 32공 트레이(53cm⨯28cm⨯6.4cm), 접수는 162공 트레이(54cm⨯28cm⨯4.6cm)에 원예용 상토(Eoullim, Shinsung Mineral Co., Korea)를 충진한 후 종자를 파종하였으며, 파종 후 12일째 본엽이 출현했을 때 접목을 실시하였다.
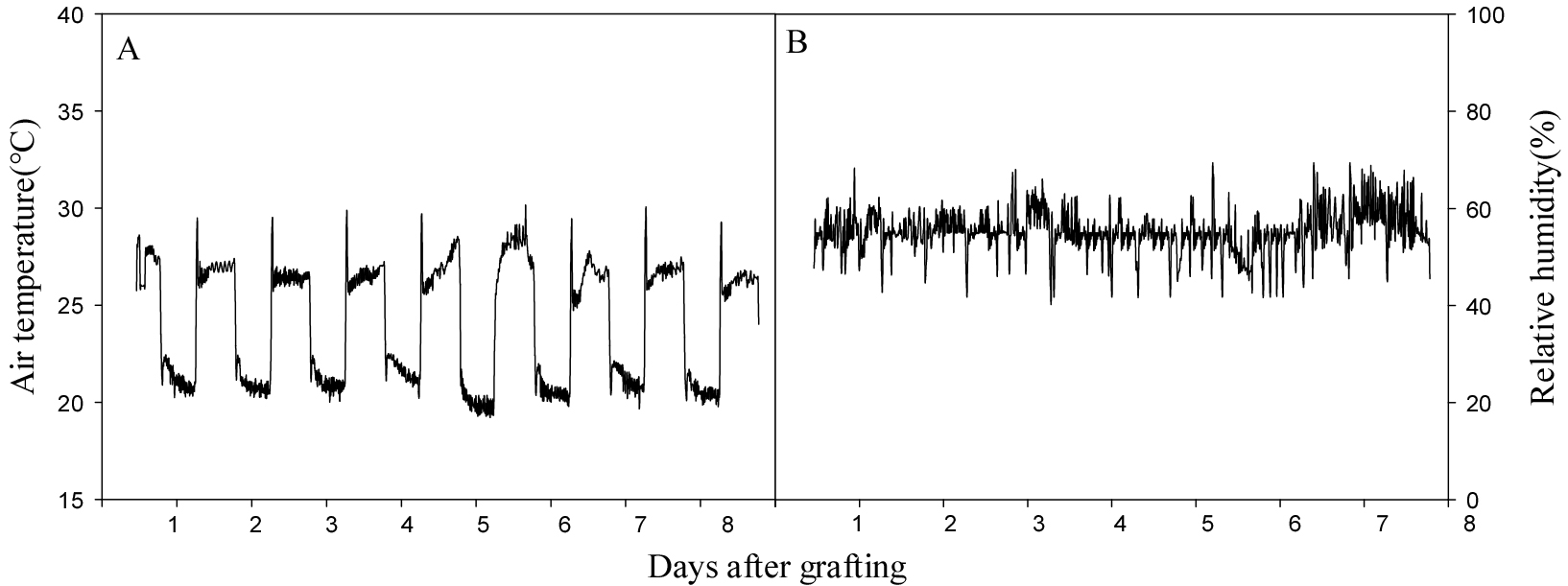
접목 후 6일간 접목 활착실에서 활착을 유도하였고 활착실의 환경 조건은 광도 10-30µmol·m-2·s-1, 온도 28°C, 상대습도 90 ± 5%로 설정하였다. 활착된 접목묘는 국립목포대학교 부속농장(34.94N, 126.67E)에 위치한 식물공장형 육묘시스템에서 생육시켰으며, 해당 육묘시스템의 환경 조건은 주간 대기온도 26 ± 2°C, 야간 18 ± 2°C, 상대습도 90 ± 5%, 광주기는 12/12시간(명기/암기)으로 설정하여 육묘장 환경과 비슷하게 조절하였다. 온·습도 측정기기(WatchDog 1000 series Micro Stations, Spectrum Technologies Inc., USA)를 이용하여 온도 및 습도 데이터를 실시간으로 수집하였다(Fig. 1). 양액 공급은 EC 1.84 dS·m-1 및 pH 6.15로 조정된 PBG 표준 양액[KNO3 505g·t-1, NH4H2PO4 115g·t-1, Ca(NO3)2H2O 708g·t-1, MgSO4·7H2O 369g·t-1, EDTA-Fe 20g·t-1, H3BO3 3g·t-1, MnSO4·4H2O 2g·t-1, ZnSO4·5H2O 0.22g·t-1, CuSO4· 5H2O 0.05g·t-1, Na2MoO4·2H2O 0.02g·t-1]을 사용하였으며, 1일 1회 저면관수 하였다.
2. 광도
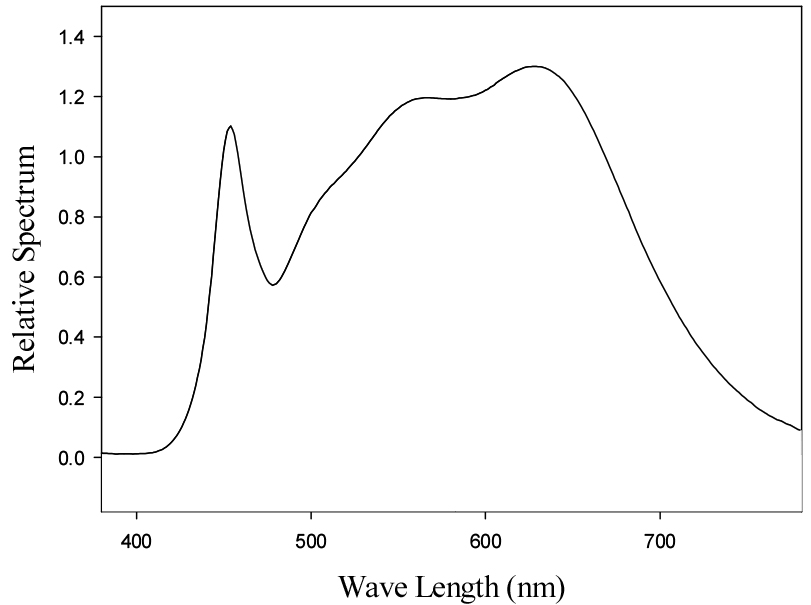
식물공장 육묘시스템에서 광원은 식물생장용 LED (FSPG- 1200OW, Shinhwa General Construction Co., Ltd., Korea)를 사용하였고(Fig. 2), 수박 접목묘에 대해 광합성유효광량자속밀도를 125 ± 25, 225 ± 25및 325 ± 25µmol·m-2·s-1로 처리하였다. 광도 처리는 활착 이후부터 21일간 지속적으로 적용하였다. 광도는 광도계(HD2101.1, Delta OHM, Italy)를 이용하여 식물체의 생장점 위치에서 측정하였다.
3. 생육조사 항목 및 측정방법
수박 접목묘는 처리당 7개체씩 3반복으로 접목 후 7일, 14일, 21일에 초장, 근장, 마디수, 엽수, 엽면적, 줄기 직경, 지상부 및 지하부의 생체중과 건물중을 측정하였다. 초장은 식물체의 지제부에서 생장점까지의 길이, 근장은 식물체 당 가장 긴 뿌리를 기준으로 지제부에서 뿌리의 끝까지의 길이를 측정하였다. 마디수와 엽수는 각각 식물체의 줄기에 있는 마디와 잎의 총 개수를 계산하였다. 엽면적은 줄기를 제외한 잎의 면적을 엽면적 측정기(LI-3100 AREA METER, LI-3100C, LI-COR, Inc., USA)를 사용하여 측정하였고, 줄기 직경은 디지털캘리퍼스(CD-PS, Mitutoyo, Japan)로 지제부 상단 1cm를 측정하였다. 지상부 및 지하부의 생체중과 건물중은 전자저울을 이용하여 측정하였으며, 건물중은 시료를 70°C 건조기(Upright Convection Oven, JSOF-250T, JSR, Korea)에서 168시간 이상 건조 후 측정하였다.
광도에 따른 수박 접목묘의 묘소질을 비교하기 위해 충실도(Compactness)와 엽면적비(LAR)를 다음의 계산식을 통해 산출하였다.
4. 통계 분석
통계 분석은 SPSS System (Version 29.0, IBM SPSS Inc., USA)을 이용하여 처리 간 평균 차이 검증을 위해 Duncan의 다중 검정을 실시하였다. 그래프는 SigmaPlot 프로그램(SigmaPlot 15.0, Systat Software Inc., USA)을 이용하여 작성하였다.
결과 및 고찰
식물공장 내 광도를 125 ± 25, 225 ± 25 및 325 ± 25µmol· m-2·s-1로 조절된 육묘시스템에서 21일 동안 재배한 후 초장을 조사한 결과, 접목 후 21일 시점에서 고온기 접목묘는 125 ± 25µmol·m-2·s-1 처리에서 57.9cm로 가장 길었으며(Table 1), 저온기 접목묘는 225 ± 25µmol·m-2·s-1 처리에서 41.8cm로 가장 길었다(Table 2). 이 결과는 상추와 토마토에서 광도가 100-250µmol·m-2·s-1 일 때 초장이 다른 처리에 비해 길었고, 300µmol·m-2·s-1에서 가장 짧았다는 연구 결과와 유사하였다(Park 등, 2013; Zheng 등, 2023). 광도가 부족한 환경에서 식물이 보다 많은 빛을 확보하기 위해 줄기 신장을 촉진하는 형태적 반응을 보인다(Hussain 등, 2020). 이는 식물이 여과되지 않은 햇빛에 도달하려는 전략적 적응으로, 낮은 광도에서 신장이 증가하는 본 연구 결과를 뒷받침한다. 그러나 과도한 줄기 신장은 식물의 쓰러짐이나 기계적 손상을 유발하여 생육 안정성을 저하시킬 수 있다(Raza 등, 2019).
Table 1.
Growth characteristics of watermelon grafted seedlings of high-temperature season as affected by days after grafting and light intensity treatments.
| Days after grafting |
Light intensity (µmol·m-2·s-1) |
Plant height (cm) |
Root length (cm) |
No. of nodes (/plant) |
No. of leaves (/plant) |
Leaf area (cm2) |
Stem diameter (mm) | Fresh weight (g) | Dry weight (g) | |||
| Shoot | Root | Shoot | Root | |||||||||
| 7 | 125 ± 25 | 11.8 bz | 18.6 a | 3.4 b | 5.9 a | 104.9 ab | 5.9 a | 7.5 a | 2.8 a | 0.64 a | 0.11 a | |
| 225 ± 25 | 13.3 ab | 18.5 a | 3.8 ab | 6.1 a | 95.2 b | 6.0 a | 7.1 a | 3.7 a | 0.73 a | 0.12 a | ||
| 325 ± 25 | 15.0 a | 18.0 a | 4.1 a | 6.1 a | 115.8 a | 6.0 a | 7.2 a | 3.3 a | 0.66 a | 0.09 b | ||
| 14 | 125 ± 25 | 18.9 b | 16.5 a | 4.7 a | 6.1 a | 101.3 a | 6.0 a | 8.9 b | 1.6 a | 0.68 c | 0.06 a | |
| 225 ± 25 | 23.2 a | 17.2 a | 4.8 a | 6.4 a | 103.8 a | 5.8 a | 9.0 b | 1.6 a | 0.90 b | 0.06 a | ||
| 325 ± 25 | 24.0 a | 18.4 a | 5.2 a | 6.2 a | 109.4 a | 6.1 a | 10.1 a | 1.7 a | 1.13 a | 0.02 b | ||
| 21 | 125 ± 25 | 57.9 a | 19.5 a | 7.2 b | 11.9 a | 113.4 c | 6.0 b | 12.0 c | 2.9 b | 1.18 a | 0.13 a | |
| 225 ± 25 | 55.0 a | 20.1 a | 8.2 a | 11.7 a | 126.5 b | 6.4 a | 15.6 a | 3.3 a | 1.20 a | 0.14 a | ||
| 325 ± 25 | 45.5 b | 20.4 a | 7.8 ab | 11.2 a | 153.8 a | 5.9 b | 14.6 b | 2.8 b | 1.11 a | 0.10 a | ||
Table 2.
Growth characteristics of watermelon grafted seedlings of low-temperature season as affected by days after grafting and light intensity treatments.
| Days after grafting |
Light intensity (µmol·m-2·s-1) |
Plant height (cm) |
Root length (cm) |
No. of nodes (/plant) |
No. of leaves (/plant) |
Leaf area (cm2) |
Stem diameter (mm) | Fresh weight (g) | Dry weight (g) | |||
| Shoot | Root | Shoot | Root | |||||||||
| 7 | 125 ± 25 | 14.8 abz | 13.6 a | 2.7 a | 3.8 b | 70.9 b | 4.7 a | 6.3 b | 1.3 a | 0.36 b | 0.03 c | |
| 225 ± 25 | 16.2 a | 14.7 a | 2.6 a | 4.4 a | 91.8 a | 5.0 a | 7.3 a | 1.9 a | 0.51 a | 0.06 a | ||
| 325 ± 25 | 14.2 b | 13.7 a | 2.1 b | 4.6 a | 72.5 b | 5.0 a | 6.0 b | 1.1 a | 0.38 b | 0.04 b | ||
| 14 | 125 ± 25 | 22.2 a | 15.0 b | 2.9 b | 6.2 b | 76.3 b | 4.9 ab | 8.3 b | 1.3 c | 0.53 c | 0.04 c | |
| 225 ± 25 | 24.5 a | 18.3 a | 3.6 a | 6.8 a | 148.4 a | 5.1 a | 11.8 a | 2.1 a | 0.87 a | 0.08 a | ||
| 325 ± 25 | 22.3 a | 14.2 b | 3.4 a | 5.9 b | 128.5 ab | 4.8 b | 8.5 b | 1.6 b | 0.67 b | 0.06 b | ||
| 21 | 125 ± 25 | 36.3 b | 15.7 b | 5.2 a | 7.8 a | 148.1 ab | 5.3 b | 10.4 b | 1.8 b | 0.89 b | 0.08 a | |
| 225 ± 25 | 41.8 a | 18.1 a | 5.4 a | 7.5 a | 163.9 a | 5.8 a | 12.1 a | 2.2 a | 1.05 a | 0.10 a | ||
| 325 ± 25 | 34.8 b | 16.1 b | 4.4 b | 6.1 b | 132.4 b | 5.5 ab | 10.1 b | 1.8 b | 0.87 b | 0.09 a | ||
줄기 직경은 고온기 접목묘는 접목 후 21일에 225 ± 25µmol· m-2·s-1 처리에서 6.4mm로 가장 두꺼웠으며 다른 처리와 유의한 차이를 보였다. 또한, 저온기 접목묘에서도 225 ± 25µmol· m-2·s-1 처리에서 줄기 직경이 5.8mm으로 가장 두꺼웠다. 이 결과는 상추와 토마토를 대상으로 50-450µmol·m-2·s-1의 광도 처리를 한 실험에서도 두 작물 모두 250 µmol·m-2·s-1에서 줄기 직경이 가장 두꺼웠다는 연구 결과와 일치하였다(Chen 등, 2022; Zheng 등, 2023). 일반적으로 광도가 줄어들면 식물은 광 포집을 극대화하기 위해 줄기 신장을 우선적으로 촉진하지만, 이 과정에서 줄기 직경은 감소하는 경향이 있다(Gao 등, 2019; Tang 등, 2022). 이는 낮은 광도에서 탄수화물의 우선적 분배가 줄기 신장에 집중되며, 줄기 직경 발달에 충분한 에너지가 사용되지 않기 때문으로 해석된다(Wu 등, 2017). 본 연구에서도 낮은 광도에서 초장이 증가한 반면, 줄기 직경은 감소하는 경향을 보였는데, 이는 기존 연구에서 보고된 낮은 광도에서의 줄기 직경 감소 현상과 일치한다. 줄기 직경 발달은 식물의 구조적 안정성과도 직결되는데 농업 생산에서 낮은 광도는 식물 높이를 증가시키고 줄기 직경을 감소시켜 쓰러짐의 위험을 증가시키며, 이는 영양분, 수분, 광합성 산물의 운반을 방해하여 생산성 감소로 이어질 수 있다(Wu 등, 2016). 따라서, 작물 생육 최적화를 위해 광을 적절히 조절하여 줄기 신장을 억제하면서도 충분한 줄기 직경 발달을 유도하는 것이 중요하다.
근장은 초장과는 반대로 고온기 접목묘는 접목 후 21일에 325 ± 25µmol·m-2·s-1 처리에서 20.4cm로 가장 길었고, 저온기 접목묘에서 접목 후 14 및 21일에 225 ± 25µmol·m-2·s-1 처리에서 18.3 및 18.1cm로 가장 길었다. 이러한 결과는 상추가 200-250µmol·m-2·s-1 조건에서 근장이 가장 길었다는 연구 결과와 일치하였다(Park 등, 2013; Chen 등, 2022). 광도는 뿌리 발달에 중요한 영향을 미치며, 이는 빛에 의해 조절되는 옥신 신호전달 기작과 밀접한 관련이 있다(Swarup 등, 2008). 빛은 어린 잎에서 옥신 합성을 유도하며, 이 옥신은 극성 수송을 통해 뿌리로 이동하여 주근과 측근 발달을 촉진시킨다(Salisbury 등, 2007). 본 연구에서 저온기 접목묘의 경우 225 ± 25µmol·m-2·s-1에서 근장이 가장 길었는데, 이는 적절한 광도가 옥신의 생합성과 이동을 조절하여 뿌리 성장을 촉진했을 가능성을 시사한다.
마디수는 접목 후 21일째에 225 ± 25µmol·m-2·s-1 처리에서 고온기 접목묘는 8.2개, 저온기 접목묘는 5.4개로 마디수가 가장 많았다. 이는 줄기 신장이 촉진되면서 초장이 길어졌고, 그에 따라 마디수 또한 증가한 것으로 판단된다.
엽수는 고온기 접목묘에서 모든 처리 간 유의한 차이가 없었으나, 저온기 접목묘는 접목 후 21일에 125 ± 25, 225 ± 25 µmol·m-2·s-1 처리에서 각각 7.8, 7.5개로 엽수가 가장 많았다. 이러한 결과는 상추의 엽수가 100, 200µmol·m-2·s-1 조건에서 많았다는 연구 결과와 유사하였다(Park 등, 2013). 엽수는 광합성 활성, 탄소 동화작용 및 생장호르몬 분포 등에 영향을 받으며 광도는 이러한 생리적 과정을 조절하는 중요한 요소로 작용한다(Taiz와 Zeiger, 2010).
엽면적은 고온기 접목묘에서 접목 후 21일에 325 ± 25µmol· m-2·s-1 처리에서 153.8cm2로 가장 넓었다. 또한, 저온기 접목묘는 접목 후 7, 14, 21일 모두 225 ± 25µmol·m-2·s-1 처리에서 각각 91.8, 148.4, 163.9cm2로 가장 넓었다. Jang 등 (2014) 에 따르면 오이 접목묘에 조사되는 광도가 증가할수록 엽면적이 증가하는 경향을 보이는데 본 연구의 고온기 접목묘의 엽면적 증가 경향과 유사한 결과를 나타낸다(Jang 등, 2014). 이는 낮은 광도에서 식물이 줄기의 신장을 촉진시키기 위해 에너지를 사용함으로써 잎의 발달을 저해한 것으로 사료된다(Morgan 등, 2002). 반면 높은 광도에서 식물이 줄기 신장보다 엽면적을 확대하여 광합성을 최적화한 것으로 사료된다. 이러한 결과는 낮은 광도에서 식물이 제한된 광을 최대한 활용하기 위해 초장을 늘려 빛에 더 잘 접근할 수 있도록 하는 반면 이는 엽면적 감소로 이어질 수 있음을 시사한다.
지상부 및 지하부의 생체중은 고온기 접목묘에서 접목 후 21일에 225 ± 25 µmol·m-2·s-1 처리에서 각 15.6 및 3.3g, 저온기 접목묘도 12.1 및 2.2g으로 가장 높았다. 또한, 지상부 및 지하부 건물중 역시 고온기 접목묘는 지상부 1.20g, 지하부 0.14g, 저온기 접목묘는 지상부 1.05g, 지하부 0.10g으로 225 ± 25 µmol·m-2·s-1에서 가장 높았지만 유의한 차이는 나타나지 않았다. 이러한 결과는 상추에서도 250µmol·m-2·s-1에서 지상부와 지하부의 생체중과 건물중이 가장 높았다는 연구 결과와 유사한 경향을 보였다(Chen 등, 2023). 광도는 식물의 광합성 효율과 생체량 축적에 중요한 영향을 미친다(Zhang 등, 2015). 일반적으로 낮은 광도에서 탄소 광합성 동화 효소의 활성이
감소하고, 이는 광계 II(PSII) 광화학의 유효 양자 수율을 감소시켜 광합성 효율 저하로 이어진다(Zheng 등, 2011; Branco 등, 2017). 따라서 낮은 광도에서 자란 식물은 낮은 광합성 효율로 인해 생체중과 건물중이 감소하는 경향을 보인다(Pires 등, 2011; Fu 등, 2012; Branco 등, 2017). 본 연구에서도 100-150µmol·m-2·s-1에서 생체중과 건물중이 상대적으로 낮았으며, 이는 광합성 제한으로 인해 동화산물 축적이 감소했기 때문일 가능성이 있다.
충실도와 엽면적비(LAR)는 묘의 소질을 평가하는 중요한 생육 지표로 높을수록 강건한 묘로 간주된다(Chen 등, 2023). 본 연구에서 고온기 접목묘와 저온기 접목묘 모두 광도 증가에 따라 충실도가 향상되었으며 특히 고온기 접목묘에서 이러한 경향이 뚜렷하게 나타났다(Figs. 3 and 4). LAR의 경우 고온기 접목묘에서 광도가 높을수록 유의하게 증가하였고 이는 높은 광도 조건에서 지상부 건물중 대비 엽면적이 효과적으로 발달했을 것으로 판단된다. 반면, 저온기 접목묘에서 처리 간 유의한 차이가 없어 광도 변화에 대한 민감도가 낮거나 다른 요인들이 생장에 더 큰 영향을 주었을 것으로 판단된다.
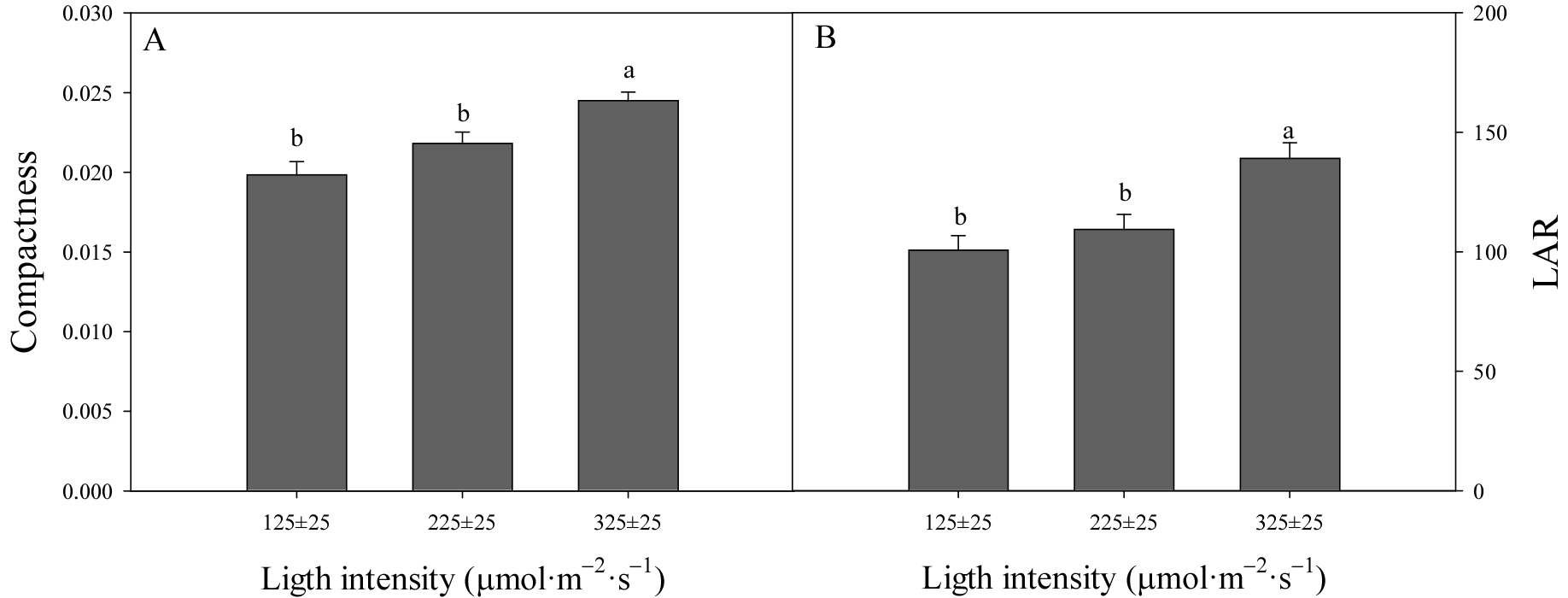
Fig. 3.
Compactness (A) and leaf area ratio (LAR) (B) of watermelon seedlings grafted with ‘BravoKkul’ scion onto ‘BullojangSaeng’ rootstock, measured at 21 days after grafting under different light intensities. Vertical bars represent the standard error of the mean. Different letters indicate significant differences based on Duncan’s multiple range test at p < 0.05.
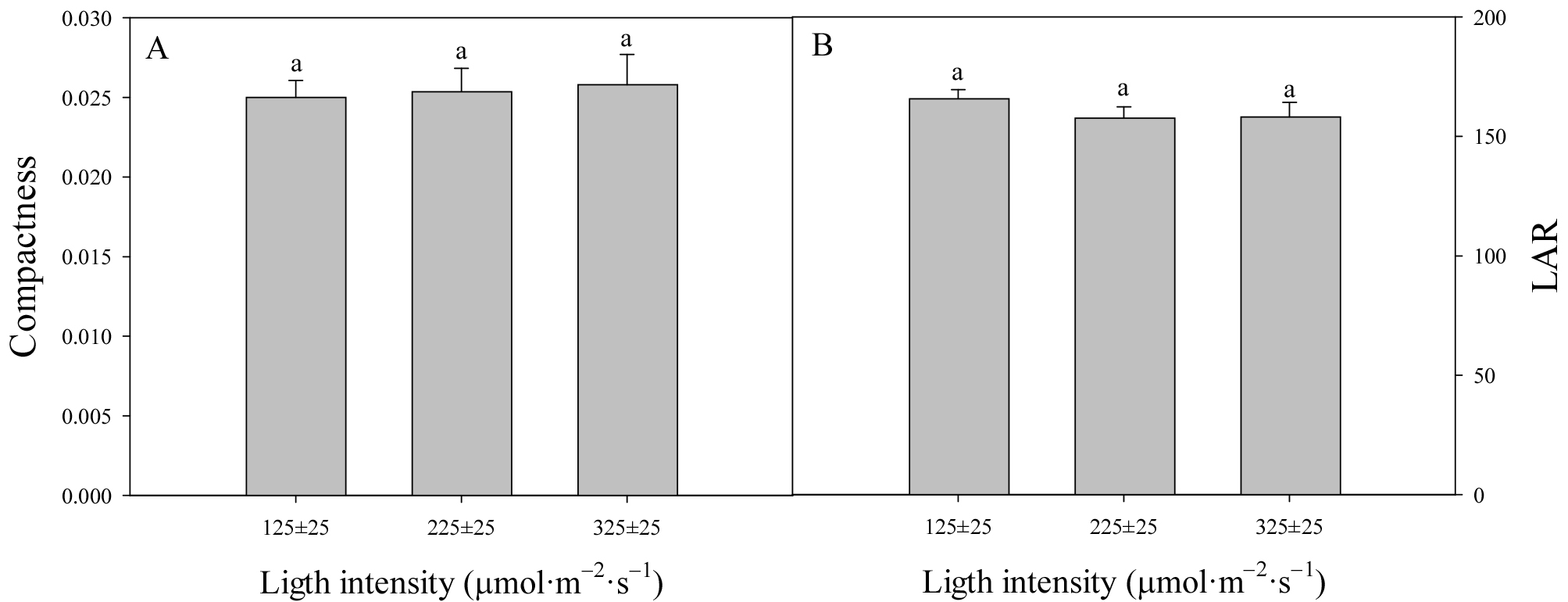
Fig. 4.
Compactness (A) and leaf area ratio (LAR) (B) of watermelon seedlings grafted with ‘SantaKkul’ scion onto ‘DongjangGun’ rootstock, measured at 21 days after grafting under different light intensities. Vertical bars represent the standard error of the mean. Different letters indicate significant differences based on Duncan’s multiple range test at p < 0.05.
본 연구 결과 수박 접목묘의 생육 특성은 전반적으로 225 ± 25µmol·m-2·s-1에서 높은 값을 보였으나 묘소질을 고려했을 때 325 ± 25µmol·m-2·s-1에서 더욱 강건한 묘가 형성된 것으로 판단된다. 따라서, 식물공장에서 접목묘의 생육 최적화를 위해서 단순한 생육 수치뿐만 아니라 묘의 품질을 종합적으로 고려하여 적절한 광도를 설정하는 것이 중요하며 325 ± 25 µmol·m-2·s-1이 보다 안정적인 생육을 유도할 수 있을 것으로 판단된다.
