

서 론
발광 다이오드(LEDs)는 제조기술의 발달과 가격하락 의 영향으로 많은 분야에서 더욱더 보편화되고 있다. 이 러한 이유로는 상대적으로 높은 전기에 대한 광 에너지 의 변환 효율과 다양한 스펙트럼, 상대적으로 낮은 표면 온도, 긴 수명, 가스가 없고 경도가 높은 재질 특성 때 문이다. 특히, 흰색 LED 칩은 그 효율(lumen/W)이 2010년에 75, 2016년 150, 2020년에는 200에 도달 할 것으로 기대된다(US Department of Energy, 2011).
광은 일반적으로 광강도, 광질, 광주기라는 요소들이 식물의 생육, 형태형성, 2차 대사산물의 생성(Lee et al., 2016) 등에 관여한다. 이러한 광의 3가지 요소들은 식물 의 생육과 발달에 중요한 환경요인으로써 식물의 생리학 적, 형태형성학적, 해부학적으로 뿐만 아니라 광합성의 중요한 에너지원으로서 그 역할을 수행한다(Taiz and Zeiger, 1991). 이러한 광의 요소 중에서 광질은 광수용 체와 관련이 있는데, 파이토크롬(phytochromes; 적색과 근적외선), 크립토크롭(cryptochromes; 청색과 UV-A), 포 토트로핀(청색과 UV-A), UVR8 (UV-B) 등이 현재까지 보고되었다(Son et al., 2016). 이러한 광수용체들은 다양 한 신호 전달 경로를 통해서 광형태형성과 식물의 생장 과 발달을 유도하며(Carvalho et al., 2011; Park et al., 2013) 특히 가시광선 영역 중 적색과 청색광 파장은 엽록 소에 의해서 가장 효과적으로 흡수되는 파장이며 광합성 에 있어 매우 중요한 에너지원으로써 요구된다(McCree, 1972). 적색광(620-670nm)은 파이토크롬을 활성화시켜 종자발아, 착과, 꽃눈 분화를 조절할 수 있으며, 청색광 (420-480nm)은 크립토크롬이나 포토트로핀을 활성화시 켜 형태형성, 꽃눈의 형성, 굴광성, 엽록체의 운동, 기공 열림 등에 관여한다고 알려져 있다. 이러한 두 파장 영역 의 광은 식물의 생장, 발달, 형태형성에 매우 중요하게 인 식되어 식물공장이나 수직농장과 같은 식물 생산시스템에 많이 채택되고 있는 중요한 광원이다. 최근에 녹색광원에 대한 연구가 진행되어 Folta and Maruhnich(2007)는 녹색 광이 광합성의 에너지원과 시그널 작용에 유효하지 않다 고 보고하였으나, 녹색광이 식물의 생장에 유효한 에너지 원으로 작용한다는 보고도 있다(Johkan et al., 2012; Son et al., 2012). 녹색광에 대한 광합성 및 생장과 관 련한 효율성 논란은 매우 오래 지속되어 왔으며 배경광 원의 조건에 따라서 달라지는 경향을 나타내었다. 현재, 인공광 이용형 식물공장의 광원으로 형광등과 LED (RB, RGB, White) 등의 광원이 주로 사용되고 있는바, 상추 3품종에 대한 형광등과 RGB 비율에 대한 최적 광 조건에 대한 평가가 필요하다.
LED 광원의 경우 펄스폭 변조 방법(pulse width modulation)을 사용하게 되면 LED의 주파수범위(Hz)와 듀티비(전류가 흐르지 않은 시간에 대한 흐른 시간의 비 율)을 제어할 수 있기 때문에 LED와 같은 다이오드 형 태의 인공광원을 이용한 식물 생산시스템의 경우 이러한 주파수변조방식으로 광조사를 할 수 있다(Loo et al., 2009). 이러한 펄스광 조사를 할 경우 생육의 차이를 보 이지 않으면서 소비 전력량을 줄일 수 있고(Tamulaitis et al., 2005), 또한 10μs의 특정 진동수(Hz)에서 상추의 생육이 증가되는 보고가 있었다(Yoneda and Mori, 2004). 그러나, Son et al.(2016)은 10kHz과 30kHz 처리 구에서 연속광과 75% 듀티비 처리는 상추 생육에 있어 유의적 차이를 명확히 보인 반면, 1kHz 에서 75% 듀티 비 조건과 연속광 처리에서 생육시킨 상추의 생체중이 유의적인 차이가 없다고 보고하였다. 또한, 펄스광조사를 하면 식물의 순광합성은 듀티비 비율에 정의 상관관계를 보인다고 하였다(Hashimoto et al., 1987). 연속광 조건 과 비교하여 소모전력은 감소되지만, PPFD 값은 낮아지 며, 결국, 지상부 바이오매스의 증가는 이루어지지 않았 다. 그러나, 펄스광 조사는 진동수와 듀티비에 따라서 달라질 수 있는데, 75%의 듀티비는 연속광에 가까운 특 징을 나타내기 때문에, 펼스광에 대한 명확한 효과를 보 기 위해서는 50%의 듀티비 처리에서 같은 PPFD 조건 의 연속광과 펄스광의 차이를 살펴보는 것이 매우 중요 하다고 할 수 있다.
본 연구의 목적은 1)녹색광원을 전체 광원의 10-30% 조건으로 고정시키고 적색광과 청색광의 RGB비율을 변 화시켜 상추 3개 품종의 생육과 형태형성에 미치는 영 향과 2)적청색 LED 광원 기반의(4:1 비율-소비전압) 듀 티비 50% 조건에서 다양한 펄스광 조사가 상추 3품종 의 생육과 형태형성에 미치는 영향을 평가하기 위하여 수행되었다.
재료 및 방법
1. 식물재료
상추(Lactuca sativar L. cv. ‘적축면’, ‘롤로롯사’, ‘그 랜드래피드’)의 3개 품종(Asia Seed Co. Ltd., Seoul, Korea)의 종자 240립을 각각 240공 암면 파종판(한국 UR암면, Ansung, Korea)에 파종하였다. 이러한 파종판 은 형광등 광원의 광강도 120μmol·m-2·s-1, 온도 22±2°C, 습도 50-70% 조건의 재배실에 정치하고 2일 간격의 저 면관수 방식으로 관수하였다. 배양액은 야마자키 상추 배양액(NO3-N 6me·L-1, NH4-N 0.5me·L-1, Ca 2me·L-1, PO4-P 1.5me·L-1, Mg 1me·L-1, SO4-S 1me·L-1, K 4me·L-1)의 1/2 농도로 본엽이 발생한 시점부터 급액하 였다. 본엽 4매가 출현한 파종 후 21일째 각각의 묘는 같은 크기의 생육정도를 판단하여 27주를 선발하였으며 스티로폼 베드(600 × 600) 2개에 15cm의 주간 간격으로 각각의 품종 9주씩 총 27주를 정식하였다. 묘가 식재된 정식판은 박막수경장치가 설치된 3단 재배장치에 각각 정치하였으며 이러한 재배장치는 총 4대가 설치되어 실 험에 이용되었다. 양액은 양액탱크에(300L) 수중펌프 (IP217, 윌로펌프, 부산, 대한민국)를 설치하여 최상단 재 배베드로 양액을 공급하고 배수되는 양액은 아래 재배베 드로 흘려보내 최종적으로 하단의 양액탱크로 순환시켰 다. 재배베드는 PVC재질을 이용하여 성형가공하였으며 600mm(폭)과 1200mm(길이)로 제작되었으며, 가운데 압 축 스티로폼으로 공간을 나누었다. 정식 후 야마자키 상 추배양액의 EC 농도 1.2dS·m-1를 초기 농도로 설정하여 사용하였으며 2주 후에 새로운 양액으로 갱신하였고 실 험중에 pH는 조절하지 않았다.
2. LED 광조사 장치
<Experiment 1> RGB 비율에 따른 상추 생육에 대한 실험을 위해서 Red (UR101, 서울반도체, 대한민국; peak wavelength: 660nm), Green (GR101, 서울반도체, 대한민국; peak wavelength: 570nm), Blue (NB104, 서 울반도체, 대한민국; peak wavelength: 460nm) 칩을 이 용하여 각각의 광원을 독립적으로 제어할 수 있는 엣지 타입의 면광원[300(길이) × 300(높이)]을 개발하였다(Fig. 1, RIST-포항산업과학연구원, 포항, 대한민국). RGB 각 광원의 1개의 모듈은 입력전압을 1-10단계의 등간격으로 디밍 조절하였다. 이러한 광원모듈을 2줄로 4개씩 총 8 개를 식물재배상 위에 고정하여 RGB 비율을 6:3:1, 5:2.5:2.5, 3:3:4, 2:2:6, 1:1:8의 비율로 5개의 LED처리구 를 만들었으며 1개는 형광등(FHF32 SSEX-D, 씨티전기, 서울, 대한민국)을 이용하여 대조구로 설정하였다. RGB 의 비율에 따른 각 광원의 광합성유효광량자속밀도 (PPFD: photosynthetic photon flux density)는 광량자센 서(LI-190, LI-COR, Lincolin, NE, USA)와 모니터링 장 치(LI-250, LI-COR, Lincolin, NE, USA)를 이용하여 측 정하였으며, 모든 LED와 형광등의 스펙트럼 분포는 분광 방사측정기(MS-720; Eko Instruments Co., Tokyo, Japan) 를 이용하여 측정하였다(Fig. 2). 식물 재배베드 바닥 16 곳에서 모든 광처리의 PPFD값의 평균은 110μmol·m-2·s-1 이었으며, 일장은 16:8h(주:야) 조건이었다(Table 1).
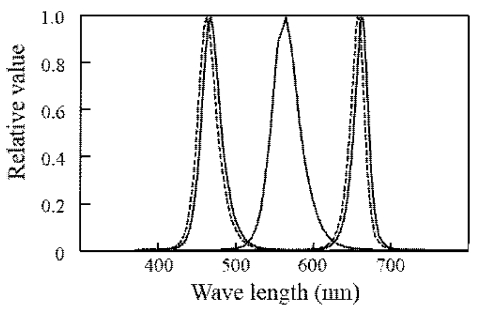
Table 1.
Mean PPFD values of each red, green, and blue LED according to RGB ratio at 9 points of the growth bed bottom and total PPFDs of each treatment.
| Light parameter | RGB ratioz | |||||
|---|---|---|---|---|---|---|
| 6:3:1 | 5:2.5:2.5 | 3:3:4 | 2:2:6 | 1:1:8 | FLy | |
| PPFDx (µmol·m-2·s-1) | ||||||
| PPFD (400-800 nm) | 108.9 | 112.7 | 108.2 | 109.2 | 109.1 | 110 |
| Red (600-670 nm) | 68.2 | 59.8 | 30.3 | 21.0 | 13.8 | 27.5 |
| Green (520-570 nm) | 31.6 | 26.1 | 31.4 | 21.6 | 12.5 | 39.6 |
| Blue (430-480 nm) | 9.1 | 26.8 | 46.6 | 66.7 | 82.8 | 27.5 |
| Far-red (700-800 nm) | 15.4 | |||||
| Fractionw (%) | 100 | 100 | 100 | 100 | ||
| PPFD (400-800 nm) | 62.6 | 100 | 28.0 | 19.2 | 12.6 | 100 |
| Red (600-670 nm) | 29.0 | 53.1 | 29.0 | 19.8 | 11.4 | 25.2 |
| Green (520-570 nm) | 8.4 | 23.1 | 43.0 | 61.0 | 76.0 | 36.3 |
| Blue (430-480 nm) | 24.8 | 25.2 | ||||
| Far-red (700-800 nm) | 13.3 | |||||
<Experiment 2> 본 실험에 사용된 LED광조사 시스템 (LCSZMS-001, 파루스, Chunan, Korea)은 광조절 장치, 환경측정장치, 유틸리티로 구성되어있으며 무선 Zigbee 통신을 사용하여 제어기와 본체의 데이터 송수신을 수행 한다. 광조사 시스템에 사용된 LED는 적색(IWS-L5056- UR-K3, peak wavelength 657nm, 이츠웰, 인천, 대한민 국)과 청색(IWS-L3512-UB-K1, peak wavelength 460nm, 이츠웰, 인천, 대한민국)을 이용하였으며 적색칩과 청색 칩이 설치된 비율은 4:1(개수) 이었다(Fig. 1). 사용된 LED 광원은 주파수 변조 방법(pulse width modulation) 을 이용하여 듀티비 50%를 고정시킨 상태에서 발생 주 파수를 50, 100, 500, 1,000, 5,000, 10,000, 25,000 Hz(20, 10, 2, 1, 0.2, 0.1, 0.04ms)조건으로 발생시켰으 며 모든 주파수 처리구에서의 PPFD값은 식물 재배베드 바닥 16곳에서의 평균값으로 구하였으며 대조구로는 적 청색 LED의 연속광으로 하였으며 같은 PPFD 조건에서 실험하였다.
3. 식물 분석 및 통계
정식 4주 후, 3개 품종의 생육조사를 실시하였다. 생 체중, 엽수, 엽장, 엽폭, 근중, 최대 근장을 측정하였다. 생 체중은 수확 후 지상부를 절단하여 전자저울(HR-A, 대현 사이언스, 서울, 대한민국)을 이용하여 측정하였으며, 이후 엽장 10mm 이상의 엽을 본엽으로 간주하여 엽수를 측 정하였다. 가장 큰 엽의 엽장, 엽폭과 최대 근장은 디지 털 버니어 캘리퍼(BD-DC200P, 흥원, 서울 대한민국)를 이용하여 측정하였고, 근중의 경우 뿌리 표면에 흡착되 어 있는 수분을 실험용 티슈(킴테크, 유한킴벌리, 서울, 대한민국)를 이용하여 수분을 완벽히 제거한 후 전자저 울을 이용하여 측정하였다.
본 실험은 완전임의 배치법으로 2반복 수행되었다. 식 물 생육 측정을 위한 모든 처리구는 반복 당 9개체를 측정하였으며, 통계분석은 SAS 통계분석 프로그램 (ver. 9.2)을 이용하였다. 평균 간의 유의성 검증을 위하여 던컨 의 다중 분산분석(Duncan’s multiple range test; DMRT)를 P<0.05 조건으로 분석하였다.
결 과
1. RGB 광조사 및 펄스광 조사 균일도
RGB 비율에 따른 재배 바닥에서의 각 LED 처리구의 PPFD값은 110±2μmol·m-2·s-1 수준으로 나타났다(Table 1). 실제 작물이 정식되고 생장하면서 군락에서의 입사 광량은 초기 바닥에서의 측정값보다는 높은 값으로 유지 되었으며 수확 전 군락에서의 광강도는 230μmol·m-2·s-1 수준으로 나타났다. 입력전압의 최소값과 최대값을 10등 분하여 조절할 수 있는 소프트웨어로 처리된 RGB의 실 제 측정된 값의 비율은 –16%에서 +26%까지 차이가 났 으나 이렇게 큰 차이가 보인(이렇게 크게 나타난) 처리 구는 6:3:1과 1:1:8의 10% 비율로 처리된 광원에서 나타 났다. RGB 실험의 대조구로 사용된 형광등의 경우 약 13.3%의 Far-red 영역이 포함되었다. 주파수 변조방식에 의한 재배베드 위에서의 듀티비 50%를 가진 펄스광에 의한 PPFD 값의 분포는 72±5μmol·m-2·s-1 수준으로 나 타났다(Fig. 2).
2. RGB 비율에 따른 상추의 생육
정식 4주 후 ‘적축면’ 상추의 생체중은 RGB 비율이 6:3:1의 경우 주당 평균 생체중이 81g인 것과 비교하여 R의 비율이 높은 처리구에서 낮은 순으로 59, 58, 58 과 62g으로 유의적 차이를 나타냈다. 그러나 형광등 처리구의 생체중 66g과 비교할 때 평균값은 6:3:1의 처리구가 높았으나 유의적 차이는 나타나지 않았다 (Fig. 3). 엽장의 경우 RGB의 비율이 6:3:1의 경우 2:2:6과 1:1:8 비율 처리구와 비교하여 유의적으로 길 게 나타났다(Fig. 3). 적축면 상추 뿌리의 생체중에 있 어서 6:3:1의 비율이 5:2.5:2.5의 처리와 비교하여 유의 적으로 높게 나타났으나, 엽수, 엽폭, 최대 근장의 부 분에서는 유의적인 차이를 보이지 않았다(Fig. 3). 롤 로로사의 경우 대조구인 형광등 처리구와 RGB 1:1:8 처리구의 생체중이 각각 54g과 34g으로 형광등 처리 구에서 유의적으로 높은 값을 보였으며, 엽수에서도 형 광등 처리구는 RGB 1:1:8 처리구보다 유의적으로 높 게 나타났다(Fig. 3). 엽장과 엽폭 또한 RGB의 비율에 따라서 달라지는데, 청색의 비율이 높아질수록 엽장은 짧아지는 경향을 나타내어 엽의 모양이 둥근 형태로 나타났다. 대조구의 뿌리 생체중과 비교하여 RGB의 비 율이 3:3:4와 2:2:6의 처리구는 유의적 차이가 없었으나, 나머지 RGB 처리는 유의적으로 낮은 값을 나타내었다. 롤로로사 품종의 최대 근장을 측정한 결과 형광등처리 에서 가장 낮은 값을 보였으며 이후 RGB의 비율에서 청색광이 증가함에 따라서 최대 근장이 증가하는 경향 을 보였다. RGB 비율이 2:2:6과 1:1:8 처리구에서 다른 나머지 처리구와 비교하여 유의적으로 높은 결과를 보 였다(Fig. 3). 그랜드래피드 품종의 경우 RGB 비율이 6:3:1, 5:2.5:2.5, 3:3:4 와 2:2:6의 처리구에서 대조구인 형광등 처리구와 RGB 비율 1:1:8 처리구와 비교하여 생체중과 엽수에서 유의적으로 높은 값을 나타내었다. 엽장의 경우 RGB 비율이 5:2.5:2.5 처리구에서 가장 높게 나타났으며 이후 청색광의 증가에 비례하여(게) 엽장은 계속 작아지는 경향을 보였다. RGB 비율이 6:3:1에서 그랜드 래피드 품종의 엽폭은 가장 길었으며 이후 청색광의 비율이 증가함에 따라서 엽폭이 감소하 는 경향을 보였다. 광원에 따른 엽장과 엽폭의 감소비 율은 달랐으나, 전체적으로 청색의 비율이 증가하면서 둥근 형태의 엽형으로 나타났으며, 적축면과 롤로로사 에서 보여준 경향과 유사한 것으로 판단된다. 뿌리 생 체중의 경우 RGB 비율이 3:3:4 처리에서 유의적으로 가장 높은 값을 보였으며, 청색광 또는 적색광이 증가 할수록 뿌리 생체중이 감소하는 경향을 나타내었으며 대조구인 형광등 처리에서 가장 낮은 값을 나타내었다. 롤로로사도 유사한 경향을 보였으며, RGB의 비율에 따 른 광합성 산물이 지하부로 이동되는 전류속도의 차이 에 의해 발생되는 것으로 생각된다. 상추 3개 품종에 조사된 광의 다른 RGB 비율은 지상부 및 지하부 생육 과 엽형을 변화시켰으며, 특히 청색광의 비율이 증가할 수록 엽장은 감소하는 경향이 모든 품종에서 나타났으 며, 청색광이 80%를 차지하는 처리구에서는 생체중이 유의적으로 감소하는 경향을 나타내었다. 또한 품종에 따른 광에 대한 각각의 반응으로 상이한 형태형성을 유 도하고 광합성량과 분배율의 차이에 의해 다른 반응을 보이는 것으로 판단된다.
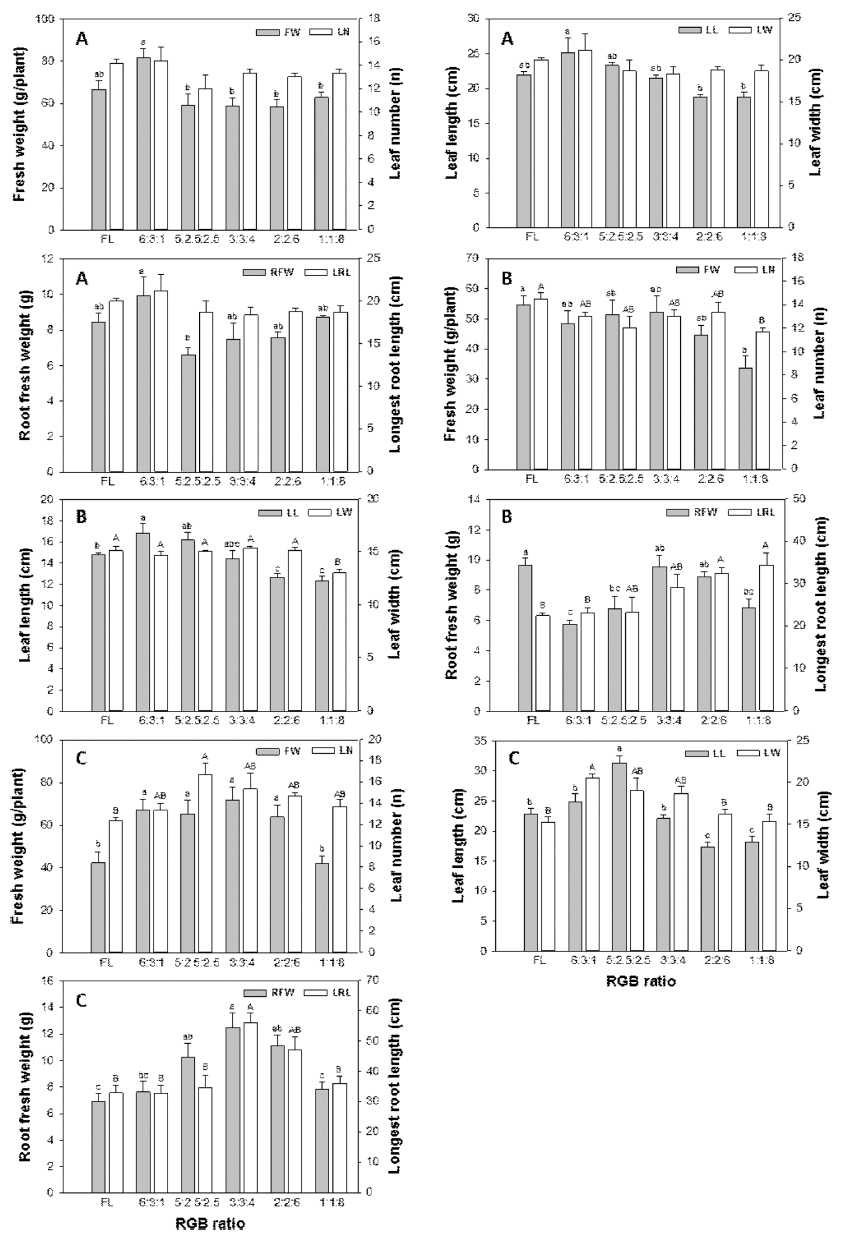
Fig. 3
Fresh weight, leaf number, leaf length, leaf width, root fresh weight, and longest root length of lettuce grown under the irradiation of the different RGB ratio (6:3:1, 5:2.5:2.5, 3:3:4, 2:2:6, and 1:1:8, relatively) with fluorescent lamp (FL) for 4 weeks after transplanting (A: Jukchukmeon, B: Lollo losa, C: Grand rapid). Data represent means and standard error (n=9). Means followed by the same letter in a graph are not significantly different by DMRT (P < 0.05).
3. 펄스광 조사
펄스광에 의한 상추 3개 품종의 엽수는 대조구인 연 속광과 비교하여 유의적으로 증가한 처리구는 없었다 (Fig. 4). 다만, 적축면 상추에서 1,000Hz에서 대조구와 비교하여 유의적으로 낮았으며 롤로로사에서는 500과 10,000Hz에서 유의적으로 낮았다(Fig. 4). 그랜드 래피 드에서는 유의적인 차이를 관찰할 수 없었다. 엽장의 경우, 대조구와 비교하여 적축면 상추의 경우 유의적 차이가 없었으나, 1,000Hz에서 낮은 값을 보였으며, 롤 로로사의 경우 유의적 차이가 발생되지 않았고, 그랜드 래피드의 경우 연속광인 대조구와 비교하여 500Hz 처 리구에서 유의적으로 크게 나타났다. 적축면 상추의 엽 폭은 대조구인 연속광과 비교하여 모든 처리구에서 유 의적 차이를 보이지 않았으나, 100Hz 처리구는 50과 1,000Hz 처리구와 비교하여 유의적으로 높게 나타났다. 롤로로사의 경우 대조구와 비교하여 5,000Hz 처리구에 서 유의적으로 적게 나타났다. 그랜드 래피드의 경우 대조구가 가장 높은 값을 보이면서 50과 5,000Hz에서 유의적으로 낮은 값을 보였다(Fig. 4). 펄스광에 의한 상추 3 품종의 생체중은 대조구인 연속광과 비교하여 모든 펄스 처리구에서 유의적으로 높지 않았다(Fig. 4). 그러나, 적축면 상추의 경우 25,000Hz 처리구는 1,000Hz 처리구와 비교하여 유의적으로 높은 값을 보였 다. 그랜드 래피드의 50Hz 처리구는 연속광인 대조구와 비교하여 유의적으로 낮은 값을 보였다. 따라서, 본 실 험에서 처리된 50에서 25,000Hz 까지의 펄스광은 같은 PPFD 조건의 연속광과 비교하여 상추의 생장에는 영향 을 주지 않는 것으로 판단되며, 일부의 처리 또는 품종 에서 형태형성 측면에서 일부 차이를 나타내었으나, 펄 스광의 추이에 따른 경향값으로 판단하기에는 어렵다고 사료된다.
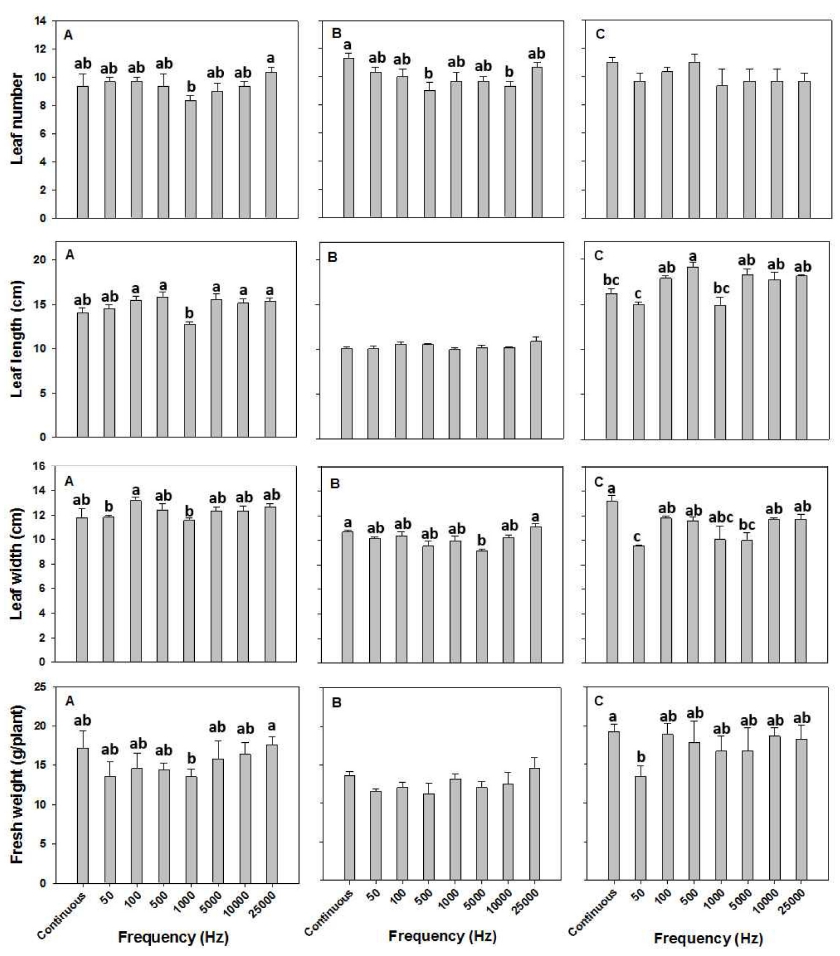
Fig. 4
Leaf number, leaf length, leaf width, and fresh weight of three cultivar lettuces (A: jukchukmeon, B: lollo rosa, C: grand rapid) grown under the different frequencies (50, 100, 500, 1,000, 5,000, 10,000, and 25,000 Hz; 20, 10, 2, 1, 0.2, 0.1, 0.04ms) of red and blue LED with 50% duty ratio using plus width modulation method for 4 weeks after transplanting. Continuous light was used with fluorescent lamp having far-red wavelength. Data represent means and standard error (n=9). Means followed by the same letter in a graph are not significantly different by DMRT (P < 0.05).
고 찰
1. RGB 비율에 따른 상추의 생육
적색광의 비율이 낮아질수록 적축면과 롤로로사의 생 체중이 감소하는 경향을 나타낸 것은 RGB 복합 또는 2 개의 광처리에서 적색광의 비율이 높은 처리구에서 낮은 처리구로 갈수록 “선망” 적축면 상추의 생체중이 유의적 으로 감소(Son and Oh, 2015) 하는 경향과 일치하였다. 상추 또는 식물의 생장에 적색광의 긍정적 효과는 여러 연구자들에 의해서 보고되었으며(Johkan et al., 2010; Kim et al., 2005; Matsuda et al., 2007), 상추를 단색광 조건과 적청 혼합광 조건에서 재배할 때 상추의 생체중 증가는 적청 혼합광에서 더욱 효과가 컸다(Son et al., 2012; Son and Oh, 2015). 또한 여러 식물에 대한 적색, 또는 청색의 단색광 보다는 혼합광원이 광합성에 매우 유리하다고 보고하였다(Goins et al., 1997; Matsuda et al., 2007; Samuolienė et al., 2011; Savvides et al., 2012). 또한 여러 파장들 중에서 적색광은 작물 생육을 증 진시키는 매우 효과적인 광파장이다(Folta and Childers, 2008). 파이토크롬에 의해서 흡수되는 적색광은 식물의 생장과 발달에 매우 중요한 역할을 하며 광합성 기구들 의 발달과 전분의 축적에 매우 중요한 에너지원으로 사 용된다(Saebo et al., 1995). 그러나, 그랜드 래피드 품종 의 경우 RGB 1:1:8을 제외한 나머지 LED 혼합 처리에 서 유의적인 차이를 보이지 않았다. 그랜드 래피드 품종 은 광질의 변화에 덜 민감한 품종으로 사료되며, RB 9:1처리구와 RGB 7:1:2 처리에서 그랜드 래피드 TBR 품종의 상추가 각각의 광처리 4주 후 생체중이 53.9와 50.34g 으로 유의적 차이가 나지 않은 결과(Son and Oh, 2015)와 유사하였다. RB광원으로만 처리된 광환경 에 G광원을 추가할 경우 상추의 바이오매스가 1.4-1.6배 증가하는 결과에 반하여, G광원이 B광원을 대체하는 역 할을 하는 것은 아니라고 보고하였다(Kim et al., 2005; Son and Oh 2015). 따라서, 녹색광원은 상추의 바이오 매스 증가에 꼭 필요한 광원으로 사료되며, 적색광원이 전체광원의 50% 이상 확보되는 조건에서 G과 B의 조 합이 필요한 것으로 판단된다.
엽채류 3품종 모두에서 엽장과 엽폭의 변화는 적색광 에서 청색광이 많아지는 광조합에서 엽의 길이가 짧아지 고, 상대적으로 엽폭은 줄어들어 엽모양이 둥근 형태에 가까워지는 것을 알 수 있다. 이러한 결과는 적색광과 청색광의 비율이 영향을 미치는데, 청색광이 증가할수록 엽이 두꺼워지면서 길이 생장이 대조구와 비교하여 억제 된다는 결과와 유사하였다(Son and Oh, 2015). 청색광 이 메소필셀 조직을 자극하기 때문에, 양자수율의 증가 와 단위파장이 갖는 에너지 증가가 엽육조직을 두껍게 한다고 하였다(Liu et al., 2014; Xiao et al., 2011). 롤 로로사와 그랜드 래피드의 지하부 생체중은 녹색과 청색 의 비율이 높았던 RGB 3:3:4에서 유의적으로 가장 높은 값을 나타냈으며, 적색광과 청색광이 증가하는 광조합으 로 갈수록 감소하는 경향을 나타내었다. 청색광의 광수용 체로 알려진 phototropins-1은 토양의 가뭄내성에 대한 자 연선택에 의해 진화적으로 형성되었다는 것을 Arabidopsis thaliana의 실험을 통하여 암시하면서 이러한 phototropins- 1은 뿌리의 생장에 직접 관련이 있다고 하였다(Galen et al., 2007). 또한 Son and Oh(2015)는 그랜드 래피드 TBR 품종의 재배 실험에서 LED를 이용한 RGB 7:2:1 처리 는 형광등과 고압나트륨 램프처리의 대조구와 비교하여 PPFD 173μmol·m-2·s-1 조건에서 지하부의 생체중이 유의 적으로 높게 나타났다고 보고하였다. 따라서, RGB의 조 합은 대상작물의 형태형성을 고려하여 둥근형태의 엽형 을 원한다면 R이 50% 이상인 조건에서 B의 비율이 높 은 조건의 광조사가 필요하며, 정상적으로 생육을 증가 시키고자 할 경우 60% 수준의 R조건과 G와 B의 비율 을 조절하는 것이 바람직하다고 판단되었다.
2. 펄스광 조사
광이용 효율(Light use efficiency)은 PPFD 값에 대한 생체중의 증가량을 나타낸 값으로 같은 PPFD 조건에서 생체중 증가에 의해 좌우 된다. 따라서 에너지 이용 효 율(Energy use efficiency)이 더 중요하다고 할 수 있다. 10과 30kHz 조건에서 50%의 듀티비를 갖는 조건은 75%의 듀티비 조건과 비교하여 에너지 이용효율은 감소 한다고 보고 하였다(Son and Oh, 2015). 그러나, 듀티비 값이 증가하면 실제 PPFD 값도 증가하기 때문에, 작물 생육에 직접적으로 영향을 미친다고 볼 수 있다. 그러나 1kHz 조건에서 듀티비 70-80%로 광조사할 경우 밀의 생장은 연속광과 차이가 없다고 보고하였다(Dong et al., 2015). 그러나, 180과 720Hz 조건의 50% 듀티비를 이 용한 LED 광조사는 대조구인 연속광과 비교하여 감자 유묘의 생장을 더욱 증가시켰으며, 또한 1kHz 이하의 낮은 주파수 영역의 듀티비 75%는 순광합성량이 연속광 과 비교하여 유의적 차이가 발생되지 않았고, 상추의 생 장에 긍정적 영향을 끼친 결과 이러한 주파수의 특징은 식물의 생장에 영향을 줄 수 있는 인자라고 평가하였다 (Son and Oh et al., 2015). 그러나, Hashimoto(1987)는 펄스광과 연속광이 같은 PPFD 조건으로 조사되었을 경 우 순광합성은 변화하지 않으며, 듀티비의 증가는 광합 성의 증가로 이어지는 양의 상관관계에 있다고 보고하였 다. 또한, 0.1, 0.2, 0.4, 0.8, 1.6, 3.2, 6.4, 12.8kHz 조건 의 펄스광 LED 조건에서 연속광조건과 비교하여 로메 인 상추의 순광합성 속도는 유의적 차이가 발생되지 않 았다(Jishi et al., 2011). Olvera-Gonzalez et al.(2013a, 2013b)는 0.1과 1Hz 조건과 50%의 듀티비 조건에서 연 속광과 비교하여 식물의 광계II (ϕPSII)의 양자수율에서 더 좋은 결과를 나타내었으며, 이러한 결과는 광합성에 긍정적으로 작용하였고, 100과 1,000Hz를 추가한 실험 에서도 광합성 형광과 관련된 인자(Fv′/Fm′, NPQ, ϕPSII, ETR, CO2흡수율)들이 연속광과 비교하여 높았고 이러한 결과 에너지를 절약하면서 광합성 효율을 증가시 킬 수 있는 광조사 방식이라고 보고하였다.
이러한 결과로 보아, 듀티비와 주파수 조건은 분명히 식물체의 생장에 영향을 줄 수 있는 인자인 것은 분명 한 것으로 사료된다. 그러나, 온도, 습도, 광과 같은 환 경요인이 식물체의 생장에 영향을 미치는 환경범위가 긍 정적 영향을 주어 효과가 상승하는 영역과 그 효과가 유지되는 영역, 과부하되어 역효과를 주는 영역으로 넓 게 정의할 수 있는 반면 식물의 광환경으로 조사되는 주파수와 듀티비의 경우 증가 또는 감소에 따른 생장의 경향이 어떠한 패턴을 가지고 있는 것은 아닌 것으로 판단된다. 또한 이 부분은 추가 실험 또는 추후 논의가 이루어져야 할 것으로 판단된다.
