

서 론
과피의 색은 포도의 품질을 결정하는 중요한 요소 중 의 하나이다. 착색계 포도의 과피 색은 안토시아닌의 함 량 및 조성에 의해서 달라지며(Castellarin과 Gaspero, 2007), 이는 품종 고유의 유전적인 특성 및 환경적인 요 인에 의해 결정된다(He 등, 2010b).
안토시아닌은 당과 배당체인 안토시아니딘의 결합으로 합성되며, 포도에는 pelargonidin(Pel), cyanidin(Cya), delphinidin(Del), peonidin(Peo), petunidin(Pet), malvidin (Mal)의 6가지의 안토시아니딘이 존재한다(Holton과 Cornish, 1995). 이 중에서 Mal이 대부분의 품종에서 가장 많은 비 율을 차지하며, Pel은 극히 적은 양으로 존재하여 일반 적으로 관찰되지 않는다(He 등, 2010a). 각 안토시아니 딘의 조성에 따라 온도, 광 조건 등의 환경적인 요인에 따른 과피 색의 안정성이 달라지며, 안토시아닌으로 합 성된 이후의 acylation 등의 추가적인 가공에 의해서도 영향을 받는다(Tamura 등, 1994).
국내에서 가장 많이 재배되고 있는 대립계 품종인 ‘거봉’은 착색이 시작되는 7-8월의 고온으로 인하여 착 색이 불량해지는 현상이 수년간 보고되고 있으며, 기후 변화로 인한 이상고온 현상의 발생 빈도가 증가하면서 앞으로 이 문제가 더 심각해질 것으로 예상되고 있다 (Lee 등, 2012). 하지만 품종 고유의 안토시아닌 조성 의 차이가 고온에 의한 착색 불량과 어떠한 상관관계 가 있는지에 대해서는 상대적으로 연구가 부족한 상황 이다.
따라서 본 연구는 착색 개시기 고온에 의한 ‘거봉’의 착색 불량에 안토시아닌의 조성이 미치는 영향을 구명하 기 위해서 수행되었으며, 비교를 위해 고온에서도 상대적 으로 착색이 잘 이루어진다고 알려진 ‘흑보석’ 포도를 함께 처리하여 분석하였다. 착색기 고온 조건에서 ‘거봉’, ‘흑보석’의 착색 반응과 시기별 두 품종의 과피의 안토 시아닌 조성을 비교하였다.
재료 및 방법
완주군 국립원예특작과학원내의 유리온실에서 포트에 재식한 2년생 ‘거봉’, ‘흑보석’ 나무를 대상으로 7월 28 일(변색기 시작, 전체 송이의 10%가 착색)부터 8월 31 일까지의 온도를 제어하여 진행하였다. 대조구는 완주 지역의 30년 평균 기온을 시간 별로 적용하였고, 고온 구는 대조구에 비해 6°C 높은 온도를 처리하였다. 품 종 별로 4주씩 처리하였고, 한 주당 4송이, 과방당 40 립씩 착과시켜 물질 분석에 활용하였다. 온도 처리가 종료된 이후 대조구와 고온구의 온도를 모두 완주지역 9월의 30년 평균 기온으로 적용하여 착색의 진행을 관 찰하였다.
과피의 개별 안토시아닌 함량을 분석하기 위해 10일 간격으로 과피 시료를 채취하였다. 과피를 과육에서 분 리하여 붙어있는 과육을 핀셋으로 최대한 제거하고 난 후, 액체질소에 얼린 상태로 막자 사발에서 분쇄하였다. 분쇄한 과피 0.1g에 MeOH와 0.1N HCl을 85:15(v:v) 로 혼합한 추출액을 2mL 넣고 암조건에서 15시간 동 안 추출한 뒤, 3,600rpm으로 10분 동안 원심분리하여 상등액을 0.45mm syringe filter에 여과하여 HPLC (1100 Series, Hewlett-Packard, Germany)로 분석하였다 . 안토시아닌 분석은 포도 재배 및 와인 국제 사무국 (International Organisation of Vine and Wine, OIV)에 서 제시한 분석 방법을 약간 수정하여 이용하였다. 안 토시아닌 분리에 이용한 컬럼은 YMC-Pack Pro C18 RS(250 × 4.6mm, 5mm, YMC, Japan)이며, 40°C에서 41분 동안 분석하였다. 안토시아닌의 함량은 518nm의 파장에서 malvidin 3-glucoside(Extrasynthese, France) 의 표준품을 이용해 작성한 표준곡선으로 정량하여 mg·g-1의 단위로 나타내었다.
모든 통계처리는 SAS Enterprise Guide 7.1 프로그램 에 의한 t-test(independent sample)로 평균간의 유의성 (P < 0.05)을 검정하였다.
결 과
변색기부터 30일 동안의 고온에 의해 ‘거봉’, ‘흑보석’ 두 품종 모두에서 과피의 안토시아닌 함량이 감소하였다 (Table 1). 수확기 과피의 안토시아닌 조성과 각각의 함 량을 비교한 결과, ‘거봉’에서 더 많은 종류의 안토시아 닌이 고온에 의해 감소하였다. 총 안토시아닌 함량은 ‘흑보석’이 대조구에 비해 26%, ‘거봉’이 71% 감소하여, ‘거봉’에서 더 크게 감소하였다. ‘거봉’에서 개별 안토시 아닌 중 가장 큰 감소율을 보이는 것은 diglucoside 및 acyl group이 있는 Mal 형태였으며, Mal-3-(trans)- coumaroyl-5-diglucoside가 가장 크게 감소하였고, 그 외에 Mal-3,5-diglucoside, Mal-3-(trans)-coumaroylglucoside 순으 로 감소하였다.
Table 1. Individual anthocyanin contents in berry skins of ‘Kyoho’ and ‘Heukboseok’ grape under control and high temperature (HT) conditions at 125 days after full bloom.
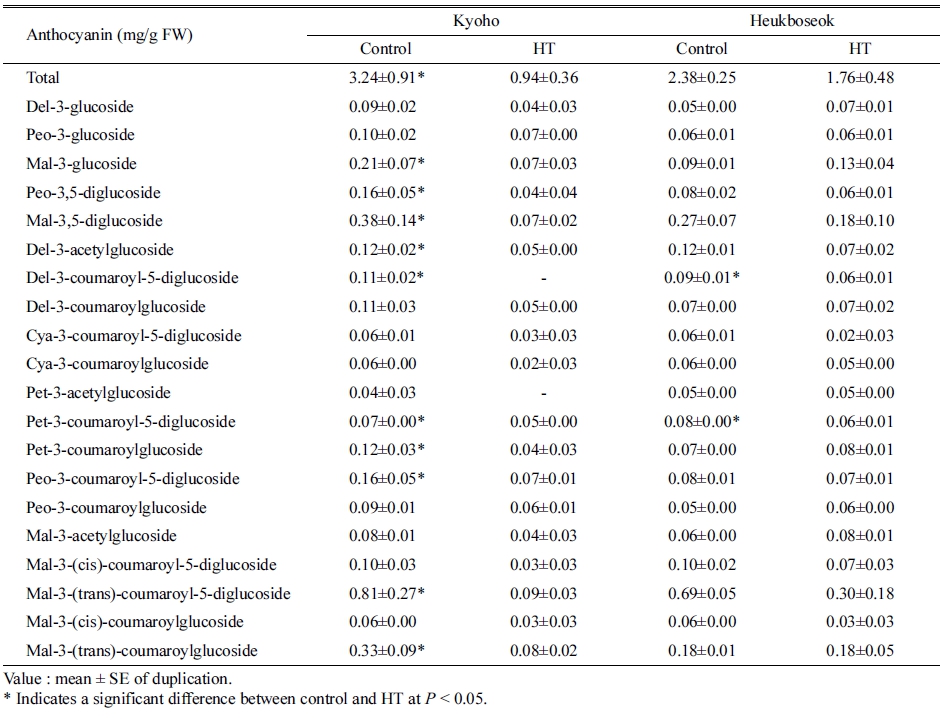 |
안토시아니딘의 그룹별로 비교하면 Mal, Del, Pet, Cya, Peo 순으로 감소하였으며, 그 중에서 Cya을 제외 한 나머지 4그룹의 함량이 대조구에 비해 유의하게 감 소하였다(Table 2). 또한, 안토시아닌의 acylation 여부나, 안토시아닌 기본 구조 중 B ring의 hydroxyl group의 개수와 관계없이 모두 고온 조건에서 함량이 감소하였다 (Table 3). Acylation이 일어난 안토시아닌(acylated)이 그 렇지 않은 형태(non-acylated)에 비해 더 크게 감소하였 고, 생합성 과정에서 B ring에 hydroxyl group이 2개 결 합되는 구조(di-hydroxylated)보다 3개가 결합되는 구조 (tri-hydroxylated)의 감소율이 더 높았다.
Table 2. Anthocyanidin contents in berry skins of ‘Kyoho’ and ‘Heukboseok’ grape under control and high temperature (HT) conditions at 125 days after full bloom.
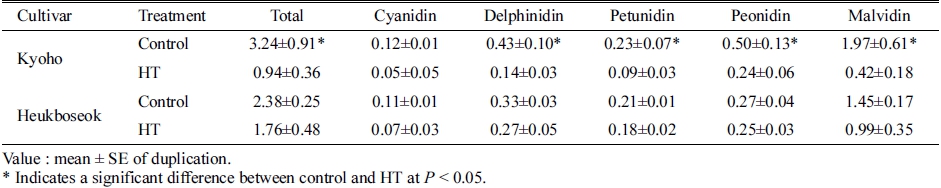 |
Table 3. Contents of different groups of anthocyanin in berry skins of ‘Kyoho’ and ‘Heukboseok’ grape under control and high temperature (HT) conditions at 125 days after full bloom.
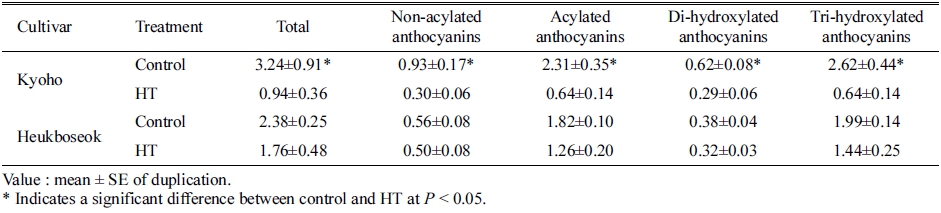 |
‘흑보석’ 과피의 총 안토시아닌은 고온 처리에 의해 감소하는 경향을 보이지만, 대조구와의 유의차가 나타나 지 않았다(Table 1). ‘흑보석’ 과피에서는 acyl group이 있는 Del, Pet-diglucoside 형태가 고온에 의해 감소하였 다. 5개의 안토시아니딘 중에서 대조구와 통계적인 차이 를 나타내는 그룹은 관찰되지 않았으며(Table 2), 안토시 아닌의 acylation 여부나, B ring의 hydroxyl group의 개 수와 관계없이 모든 그룹이 대조구에 비해 감소하지 않 았다(Table 3).
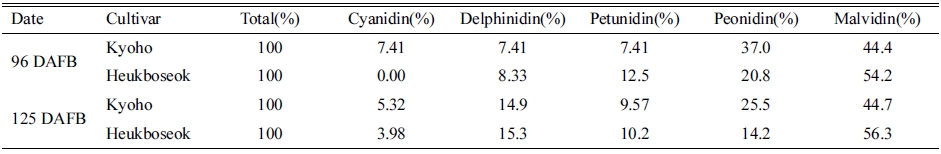
온도 처리가 종료된 후부터 수확기까지 두 품종의 안 토시아니딘의 구성 비율은 크게 달라지지 않았다(Table 4). 처리 종료 시(만개 후 96일) ‘거봉’의 과피에서는 Cya과 Peo의 비율이 흑보석에 비해 높았고, Del, Pet, Mal의 비율은 ‘흑보석’에서 더 높게 관찰되었다. 이러한 품종간 안토시아니딘의 비율의 차이가 수확기(만개 후 125일)까지 같은 경향으로 유지되었다.
Table 4. Anthocyanidin component ratios in berry skins of ‘Kyoho’ and ‘Heukboseok’ grape under high temperature (HT) conditions at 96 and 125 days after full bloom (DAFB).
 |
고온 처리에 의한 시기별 과피의 변화를 관찰한 결과, 고온 처리 2주 후부터 ‘거봉’의 착색 속도가 느려지기 시작하며 대조구에 비해 2주 이상 늦어지는 모습을 보 였다(Fig. 1). 온도 처리가 종료되고 ‘거봉’의 숙기인 8 월 하순 이후에도 더 이상 착색이 진행되지 않았다. ‘흑 보석’의 경우, 고온 처리 중에는 착색이 전혀 이루어지 지 않다가 온도 처리가 종료된 이후부터 빠르게 착색이 시작되었지만 숙기가 지나고 나서도 착색이 대조구만큼 진하게 이루어지지는 않은 것으로 조사되었다.
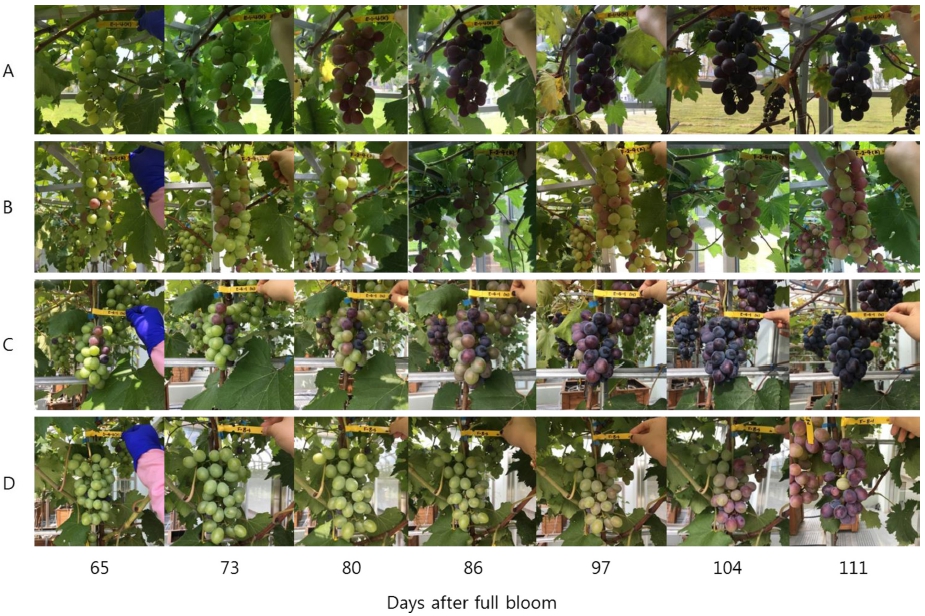
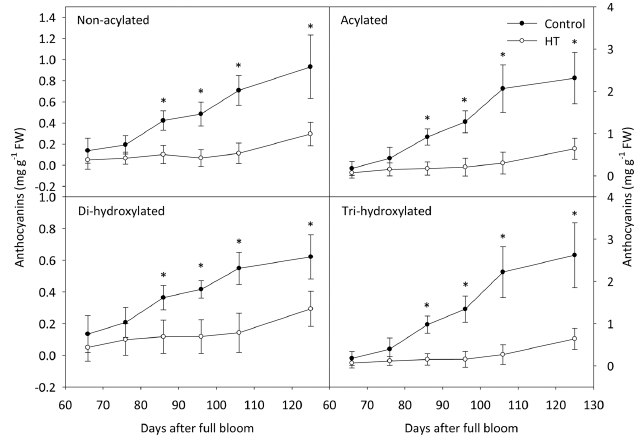
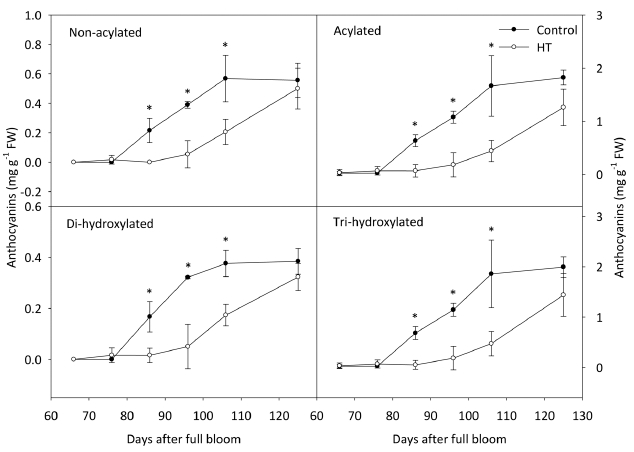
시기별 안토시아닌 함량의 변화를 관찰한 결과, ‘거봉’ 과피의 총 안토시아닌 함량은 고온 처리 20일 후부터 대조구에 비해서 낮았고 수확기까지 낮은 수준을 유지하 였다(Fig. 2). 안토시아니딘 중에서 Cya을 제외한 나머지 4그룹은 총 안토시아닌 함량과 동일한 경향을 나타내며 감소하였다. Cya의 함량은 고온에 의해 처리 30일 후부 터 감소했지만, 온도 처리의 종료 후 대조구의 수준으로 증가하였다. 안토시아닌의 형태에 따른 함량을 분석한 결과, ‘거봉’ 포도에서 acylation 및 B ring의 hydroxyl group의 개수와 관계없이 고온에 의해 처리 20일 후부 터 대조구에 비해 낮았다(Fig. 3). 고온 처리가 종료된 이후에도 안토시아닌이 축적되지 못했으며, 최종적으로 대조구의 수준만큼 증가하지 못하였다.
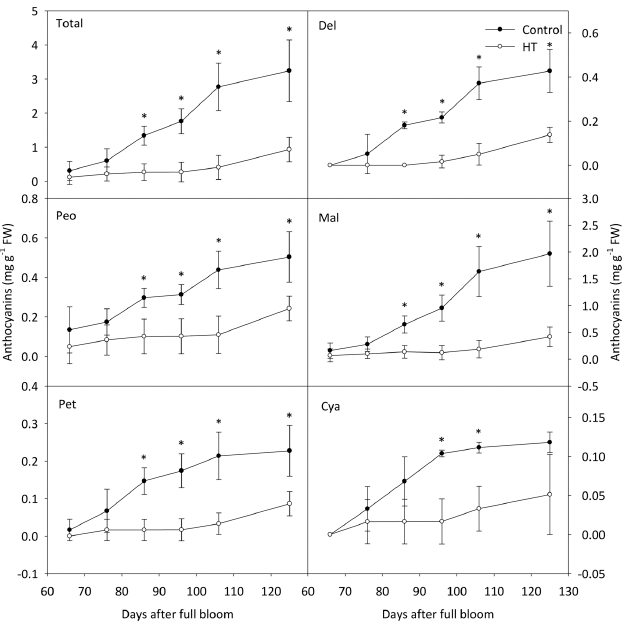
Fig. 2
Changes in anthocyanidin contents in ‘Kyoho’ grape berry skins under control and high temperature (HT) conditions. Vertical bars indicate the standard errors of the means (n = 4), and asterisks indicate significant differences using Student’s t-test at P < 0.05. Del, dephinidin; Peo, peonidin; Mal, malvidin; Pet, petunidin; Cya, cyanidin.
‘흑보석’ 과피에서는 고온 처리에 의해 안토시아닌의 합성이 억제되었지만, 온도처리가 종료된 이후 다시 축 적되기 시작하여 대조구의 수준으로 회복되었다(Fig. 4). 안토시아니딘별 함량의 변화를 분석한 결과, Cya을 제외 한 나머지 4그룹의 함량이 총 안토시아닌과 동일한 경 향을 보이며 변화하였고, 최종적으로 대조구의 함량과 차이가 나타나지 않았다. 안토시아닌의 형태에 따른 변 화를 관찰한 결과, acylation 및 B ring의 hydroxyl group의 개수와 관계없이 고온에 의해 처리 20일 후부 터 함량이 감소하였다(Fig. 5). 하지만, 온도 처리 종료 후 모든 형태의 안토시아닌이 다시 축적되기 시작하였으 며, 수확기에 대조구와 비슷한 수준으로 회복되었다.
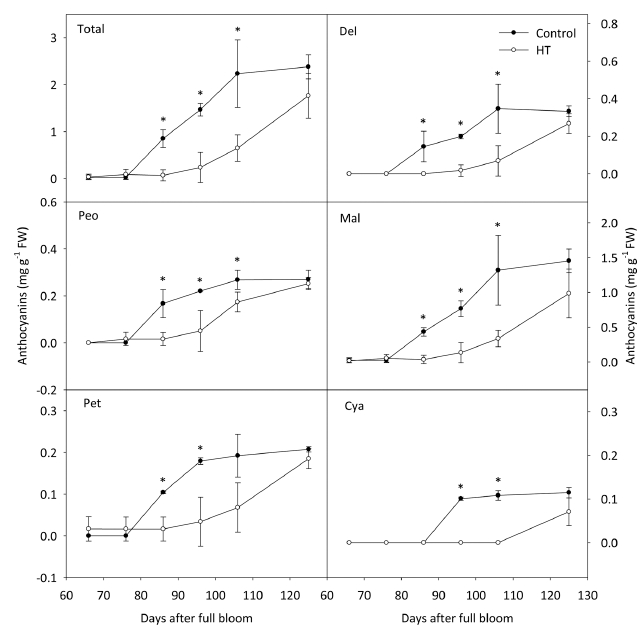
Fig. 4
Changes in anthocyanidin contents in ‘Heukboseok’ grape berry skins under control and high temperature (HT) conditions. Vertical bars indicate the standard errors of the means (n = 4), and asterisks indicate significant differences using Student’s t-test at P < 0.05. Del, dephinidin; Peo, peonidin; Mal, malvidin; Pet, petunidin; Cya, cyanidin.
고 찰
착색기의 고온에 의해 ‘거봉’과 ‘흑보석’ 두 품종 모두 의 착색이 지연되었는데(Fig. 1), 이는 주야간의 고온에 의해 포도 과피의 안토시아닌 함량이 감소하였다는 Mori 등(2004)과 Shinomiya 등(2015)의 결과와 일치하 였다. 하지만 온도 처리 종료 이후 ‘거봉’은 과피의 안 토시아닌 함량이 증가하지 못하고 착색이 정지하였지만, ‘흑보석’은 과피의 안토시아닌이 다시 축적되기 시작하 여 수확기에는 대조구의 함량과 차이가 나타나지 않았다 (Table 1). 하지만 실제 수확기의 과실은 대조구에 비해 착색이 완전히 회복되지 않은 모습을 보였다(Fig. 1). 이 는 눈에 보이는 착색의 정도가 외과피에서 넓게는 중과 피 상부까지 분포되어 있는 안토시아닌의 축적에 의해 결정되는데 비해, 안토시아닌의 정량 분석에는 외과피만 최대한 얇게 벗겨서 이용했기 때문에 이러한 차이가 나 타났다고 판단되었다. 고온에 의한 과피의 착색 불량에 서 품종에 따른 과실의 부위별 안토시아닌 축적 변화에 대해서는 추가적인 분석이 필요하다고 판단된다.
본 실험에서 ‘흑보석’ 과피의 안토시아닌 함량의 증가 가 안토시아니딘의 비율과 상관없이 전체적으로 다시 증 가하는 경향으로 나타났기 때문에(Table 4), 고온에 의한 ‘거봉’, ‘흑보석’간의 서로 다른 착색 반응은 안토시아닌 의 구성 비율이나 개별 안토시아닌의 특성 때문이 아니 라, 각 품종의 안토시아닌 생합성 과정에서 고온이 미치 는 영향이 다르기 때문으로 판단된다.
안토시아닌은 flavilium의 기본 구조를 갖고 있으며, 이는 A, B, C의 3개의 ring으로 이루어져있다(Mazza와 Francis, 1995). 포도 과피에 존재하는 안토시아니딘 그 룹은 B ring의 구조에 따라 고온 조건에서 서로 다른 축적 양상을 보인다. Di-hydroxylated 그룹(Cya, Peo)보 다 tri-hydroxylated 그룹(Del, Mal, Pet)이 고온에 더 민 감하게 반응하며 감소하며, 이는 안토시아닌 생합성 과 정에서 tri-hydroxylated 구조를 완성하는 flavonoid 3’5’- hydroxylase의 발현이 고온에 의해 감소하기 때문이다 (Mori 등, 2007). ‘거봉’ 과피에서도 고온에 의해 Mal, Del, Pet, Cya, Peo의 순서로 크게 감소하였고(Table 2), 그 중 Cya이 고온에 의해 가장 느리게 영향을 받았으며 감소율이 적었다(Fig. 2). 따라서, 진한 자주색이나 푸른 색을 나타내는 tri-hydroxylated 계열의 안토시아닌의 합 성이 고온에 의해 더 크게 억제되면서 과피의 착색이 진하게 이루어지지 못한 것이라고 판단되었다.
안토시아닌은 그 구조에 따라 온도에 대한 안정성이 달라지는데, 일반적으로 acylation이 일어나면 주 저장 기관인 액포로의 이동이 촉진되며(Gomez 등, 2009) 액 포 내에서 anthocyanic vacuolar inclusion 구조를 형성 하여(Kallam 등, 2017) 고온 조건에서 더 안정화된다고 알려져 있다(Tarara 등, 2008). 하지만, ‘거봉’ 과피에서 고온에 의해 acylation된 Mal이 가장 많이 감소하였고, non-acylated(68%)와 비교했을 때 acylated(72%)가 더 큰 비율로 감소하였다(Table 3). 기존의 연구 결과에서도 고온에 의해 non-acylated의 감소 비율이 높거나(De Rosas 등, 2017), 반대로 acylated가 더 많이 감소되는 상반된 결과가 보고되었다(Azuma 등, 2012). 본 연구의 결과를 통해, 착색 초기의 고온에 의해 과피의 착색이 불량해지는 이유는 기존에 합성된 안토시아닌이 분해되 었다기보다 전체적인 안토시아닌의 생합성이 억제되었기 때문임을 알 수 있었다. 따라서, ‘거봉’의 안토시아닌에 서 가장 많은 비율을 차지하는 acylation된 Mal의 감소 율이 가장 높았던 것으로 판단되었다.
과피에 존재하는 당은 안토시아닌의 합성 재료로서 중 요한 역할을 하며, 과실의 당이 일정 함량 이상으로 증 가하지 못하면 안토시아닌의 축적이 이루어지지 않는다 (Sadras와 Moran, 2012). 안토시아닌의 축적으로 착색이 이루어지는 사과의 경우, 야간의 고온에 의해 과실의 당 함량이 감소하여 착색이 불량해진다고 보고되었다(Ryu 등, 2015). 하지만 ‘거봉’과 ‘흑보석’의 과피 내 당 함량 은 고온에 의해 감소하지 않았으며, 거봉의 당도가 흑보 석의 당도에 비해 오히려 높은 경향을 나타내었다(데이 터 미제시). 이는 주야간 고온에 의해 착색이 억제된 경 우에 포도의 당도가 높아지거나(Shinomiya 등, 2015), 당 함량에 차이가 나타나지 않았다는 보고(Koshita 등, 2015)와 비슷한 결과로, 과피의 당 함량은 변색 초기의 고온에 의해 착색에 필요한 수준 이하로 감소하지 않는 것으로 판단되었다. 따라서, 고온 조건에서 착색이 불량 해지는 것은 당 함량의 감소 때문이 아닌, 안토시아닌 생합성과 연관된 유전자 발현과 효소의 활성이 고온에 의해 억제되었기 때문으로 생각되었다(De Rosas 등, 2017; Mori 등, 2004; Mori 등, 2007).
따라서, 본 실험의 결과를 통해 변색 초기의 고온으로 인한 ‘거봉’과 ‘흑보석’의 서로 다른 착색의 반응은 두 품 종의 안토시아닌 생합성에 고온이 미치는 영향이 다르기 때문으로 판단된다. 변색 초기의 이상 고온으로 인한 포도 의 착색 불량을 경감시키기 위해서는, 두 품종의 안토시아 닌 생합성 과정에서 차이를 가져오는 단계를 밝혀낼 필요 가 있으며, 그에 따른 후속 연구가 필요하다고 판단된다.
