

서 론
온실 내 작물의 최종 생산량은 캐노피 광합성 산물의 누적량에 의해 결정되므로(Monteith, 1965), 캐노피 광합 성은 작물의 생육과 발달을 예측하고 온실 내 CO2 시비 전략을 수립하는 데에 중요한 지표이다(Son 등, 1999;Hu 등, 2011;Jung 등, 2016). 작물의 광합성에는 광도, 온도, CO2 농도, 상대습도 등 다양한 기상 변수가 영향 을 미치며, 이는 시간에 따라 끊임없이 변화한다. 또한 엽 광합성은 기상 요인뿐 아니라 엽각, 엽 분포 등 작물 의 구조적 요인에도 영향을 받는다(Kim 등, 2016). 이러 한 구조적 요인은 일중 태양의 움직임에 따라 계속 변 화하며, 작물이 생육함에 따라 달라진다(Caldwell 등, 1986). 또한 온실 내에서의 높은 재식 밀도는 인접한 개 체 간에 상호 차광을 유발해 작물 수광에 영향을 미친 다(Chen 등, 1999). 따라서 작물의 광합성에 대한 정확 한 분석을 하기 위해서는 일중 시간에 따른 작물의 정 확한 수광량 예측이 필요하다.
그러나 작물의 수광을 정확하게 실측하는 데에는 많은 시간과 비용, 노력이 요구되기 때문에, 태양위치 변화에 따른 작물 수광을 계산할 수 있는 단순화된 모델링 접 근이 시도되어 사용되고 있다(Marcelis 등, 1998). 광합 성 계산에는 작물 엽의 광반응곡선을 기반으로 하는 직 각쌍곡선 모델이 개발되어 이용되고 있으며, 캐노피 수 광은 각 엽 광합성의 총합으로 계산된다(Acock 등, 1971;Thornley 1976;Goudriaan 등, 1995). 또한 작물 수광에 대한 용이한 계산을 위해 작물 캐노피 수직 광 분포는 상단부부터 하단부에 이르기까지 지수적으로 감 소하는 형태를 띈다고 가정한다(Monsi와 Saeki, 1953). 이러한 단순화된 접근은 매우 간편하다는 장점이 있으며 균일하고 밀집된 작물 군락에서는 높은 정확도를 보이는 반면, 불균일한 형태의 캐노피에서는 예측의 정확도가 감소한다(Vos 등, 2007).
이러한 단점에 대하여 구조-기능적 작물 모델링은 컴 퓨터를 이용한 3차원 분석을 통하여 작물 수광량을 정 확히 예측하고자 하였다. 3차원 그래픽 기술의 발달로 인해 작물의 구조를 3차원 가상 공간에 표현하는 것이 가능해졌으며(Smith, 1984;Prusinkiewicz, 1986) 작물의 3차원 모델을 이용하여 광학 시뮬레이션을 수행하면 불 균일한 형태의 캐노피에서도 정확한 작물 수광을 계산할 수 있다. 따라서 이러한 정확한 수광 분포를 기반으로 광합성의 정확한 예측이 가능해진다(Buck-Sorlin 등, 2011;Sarlikioti 등, 2011;de Visser 등, 2014;Kim 등, 2016;Jung 등, 2018). 그러므로 구조-기능적 작물 모델 링을 이용하면 정확한 수광량과 광합성 속도를 추정할 수 있다. 그러나 기존의 3차원 작물 모델은 작물의 엽, 줄기, 과실 등의 각 기관을 실제 형태에 비하여 비교적 단순화된 형태로 표현되었다. 따라서 정확한 3차원 작물 모델을 사용한다면 수광량 및 그에 따른 광합성 예측의 정확도를 향상시킬 수 있다. 3차원 스캔을 통해 정확한 작물 형태와 표면에 대한 정보를 얻고자 하였고(Paulus 등, 2014;Zhang 등, 2016), 3차원 작물 모델에 대한 연 구는 주로 정확한 작물 형태 구축 자체에 집중되었으며 수광 분석 및 구조-기능적 작물 모델링에 사용하려는 연 구는 거의 없었다(Hosoi 등, 2011; Wahabzada 등, 2015;Behmann 등, 2016;Moriondo 등, 2016). 본 연구에서는 파프리카의 3차원 스캔 모델을 사용하여 광학 시뮬레이 션을 수행하고, 엽 광합성 모델을 이용하여 특정 환경 조건 하에서 캐노피 수광 분포와 광합성 속도 분포를 예측하고자 하였다.
재료 및 방법
1. 재배조건 및 3차원 모델 구축
실험에는 함안에 위치한 국립원예특작과학원 시설원예 연구소(35.2°N, 128.4°E)의 벤로형 온실에서 파프리카 (Capsicum annuum L., cv. Scirocco)를 사용하였다. 재식 밀도는 1 m2 당 2주이며, 한 슬라브 위에 4개체씩 정식하 였다. 고해상도의 휴대용 3차원 스캐너(GO!SCAN50TM, CREAFORM, Lévis, Quebec, Canada) 를 이용하여 2017년 9월 19일에 온실 파프리카의 3차원 스캔을 진행 하였다. 3차원 스캔 결과의 정확도를 높이기 위하여 직 경 10mm의 원형 타겟을 작물의 엽, 줄기, 과실에 붙여 주었다. 3차원 스캐닝은 2mm의 해상도로 진행되었다. 3 차원 스캐닝을 통해 얻어진 스캔 메시는 스캐닝 소프트 웨어(Vxelement, CREAFORM)와 역설계 소프트웨어 (Geomagic Design X, 3D Systems, Rockhill, SC, USA) 를 이용하여 수광 시뮬레이션이 가능한 파라메트릭 모델 로 변환되었고, 이를 3차원 CAD 소프트웨어(SOLIDWORKS, Dassault Systemes, Vélizy-Villacoublay, France)에서 재 구축하였다. 작성된 3차원 파프리카 모델의 광학적 특성 을 입력하기 위해 파프리카 잎의 흡수율(Tr)과 반사율 (Ref)을 400-700nm 범위에서 spectroradiometer(BLUEWave spectrometer, StellarNet Inc., Tampa, FL, USA)를 이용하여 1nm 해상도로 측정하였다. 잎의 흡수율(Abs) 은 Abs = 1 - (Tr + Ref)로 계산되었다. 측정된 반사율과 투과율은 광학 시뮬레이션 소프트웨어(OPTISWORKS, OPTIS Inc., La Farlède, FRANCE)를 통해 3차원 파프 리카 모델에 적용해 주었다. 벤로형 온실, 암면 슬라브, 암면 배지의 3차원 모델은 실측값을 바탕으로 3차원 CAD 소프트웨어를 통해 구축하였고, 광학 시뮬레이션 소프트웨어를 통해 투과율과 반사율의 광학적 특성을 입 력해 주었다.
2. 3차원 모델 배치 및 광학 시뮬레이션
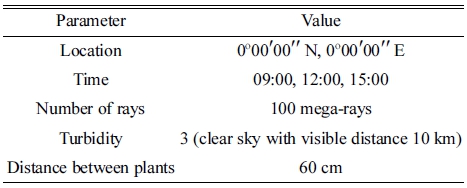
3차원 스캐닝을 통해 구축된 작물 모델을 3차원 CAD 프로그램에 배치하여 광학 시뮬레이션을 진행하였다. 주 변 개체의 유무에 따른 캐노피 수광 분포의 변화를 보 기 위해 단일 개체 배치와 9×9 정방형 배치에 대해 광 학 시뮬레이션을 각각 진행하였다. 정방형 배치는 남북 동 온실 내 작물의 재식 방향을 고려하여 V 캐노피 형 태의 두 가지가 동-서 방향을 향하도록 배치하였다. 또 한, 모델의 잎이 서로 겹치지 않는 것을 고려하여 간격 은 가로, 세로 60cm 로 설정하였고, 상호 그림자 효과를 고려하여 중심에 위치한 개체를 분석 대상으로 하였다. 광학 시뮬레이션 조건은 Table 1과 같다. 임의의 시기 2018년 4월 26일에 대하여 일중 변화를 분석하였다. 오 전 9시, 정오, 오후 3시에 대한 태양고도는 각각 43.9°, 76.4°, 42.9°이였다. 대기 혼탁도는 가시거리 10km의 일 반적인 맑은 날을 나타내는 3으로 설정하였다.
3. 광합성 속도 계산
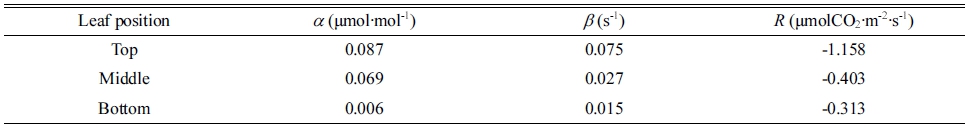
수광 시뮬레이션 결과로부터 광합성 속도를 계산하기 위해 직각쌍곡선 모델을 이용하였다. 직각쌍곡선 모델은 다음의 식으로 표현된다.
| $$P=\left(\frac{\alpha\ast I\ast\beta\ast C_i}{\alpha\ast I+\beta\ast C_i}\right)-R$$ | (1) |
P는 엽 광합성 속도(μmolCO2·m-2·s-1), I는 광합성 유효 광양자속 밀도(μmol·m-2·s-1), Ci는 세포 내 CO2 농도 (μmol·mol-1), R은 호흡 속도(μmolCO2·m-2·s-1), α는 광화 학 이용효율(μmol·mol-1), β는 카르복실화 컨덕턴스(s-1)를 의미한다(Thornley, 1974). 파프리카 엽의 상단부, 중단부, 하단부에 따른 α, β, R 값은 실측값을 바탕으로 하여 계 산하였다(Table 2). 수광 시뮬레이션은 가상의 환경에서 진행되었기 때문에 일중 변화를 고려하지 않고 온도 27°C, 상대습도 70%, 대기 중 CO2 농도 400μmol·mol-1 로 고정하였다.
결과 및 고찰
1. 캐노피 수광 분포 및 총 수광량
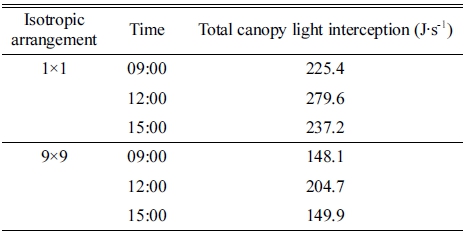
파프리카의 3차원 스캔 모델을 배치하여 수광 시뮬레 이션을 진행한 결과, 3차원 스캔 모델 표면의 수광 분포 를 계산할 수 있었다(Fig. 1). 3차원 파프리카 모델 표면 의 수광 분포는 오전 9시, 정오, 오후 3시의 태양고도에 따라 변화하였다(Figs. 1A, 1B, 1C). 작물 상단부의 수 광은 정오에 가장 높았고, 오전 9시와 오후 3시에서는 큰 차이가 없었다. 9×9 정방형 배치에 따라 주변 개체 가 늘어나면서 캐노피 수광은 감소하는 경향을 보였고, 이러한 현상은 모든 시간대에서 동일하게 나타났다(Figs. 3차원 모델 표면의 수광 분포를 높이에 따른 평균 수광 량으로 나타낸 결과 모든 시간대에서 작물 수광은 상단부 에서 하단부로 이르기까지 감소하는 경향을 보였다 (Fig. 2). 작물 최상단부의 평균 수광량은 정오에 1,979.3μmol·m-2 ·s-1로 가장 높았고, 오전 9시와 오후 3시에는 각각 1,099.2, 1,096.1μmol·m-2·s-1로 약 55% 정도의 낮은 수 광을 보였다. 특히 모든 시간대에서 1×1 배치에 비해 9×9 배치에서 수광이 급격하게 감소하였다(Figs. 2A, 2B, 2C). 수고 120cm 이상의 작물 상단부에서는 주변 개체에 의한 수광량 감소가 적었으나, 그 이하의 작물 중, 하단부에서는 수광량이 크게 감소하였다. 1×1 배치 의 경우 정오에서는 상단부에서 하단부까지 수광량이 지 수적으로 감소하는 형태를 나타냈으나(Fig. 2B), 오전 9 시와 오후 3시에는 상단부와 하단부의 수광 차이가 작 았다(Figs. 2A, 2C). 캐노피 총 수광량은 1×1 배치 시 정오에 279.6J·s-1로 가장 높은 값을 보였고, 오전 9시와 오후 3시에는 각각 225.4, 237.2J·s-1로 비슷한 값을 보였 다(Table 3). 9×9 배치 시 캐노피 총 수광량은 정오에 204.7J·s-1로 1×1 배치에 비해 약 73.2%로 감소하였고, 오전 9시와 오후 3시에는 각각 148.1, 149.9J·s-1로 1×1 배치에 비해 65.7%, 63.2%로 감소하였다.
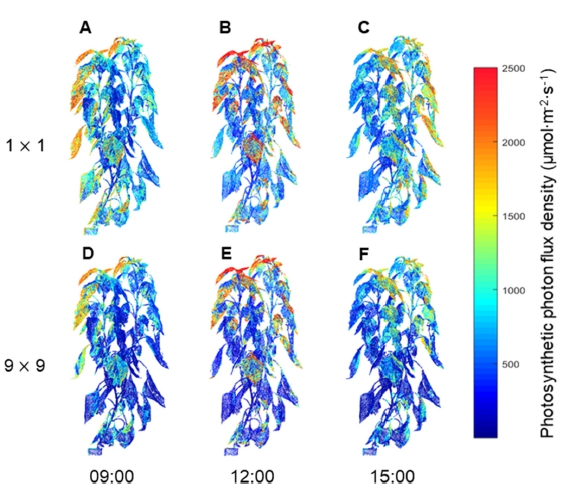
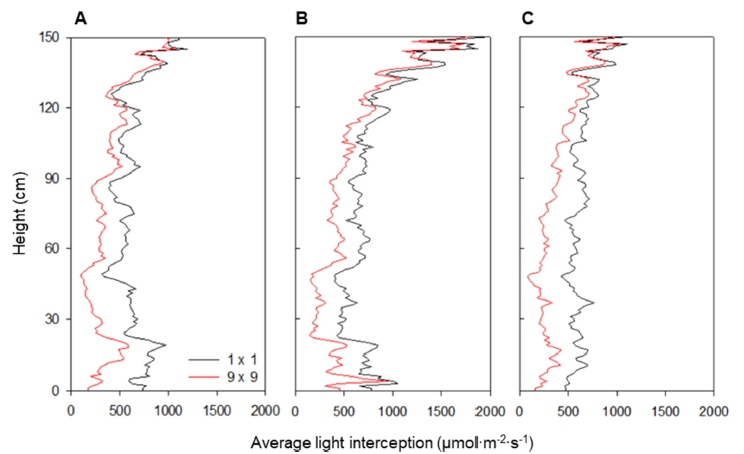
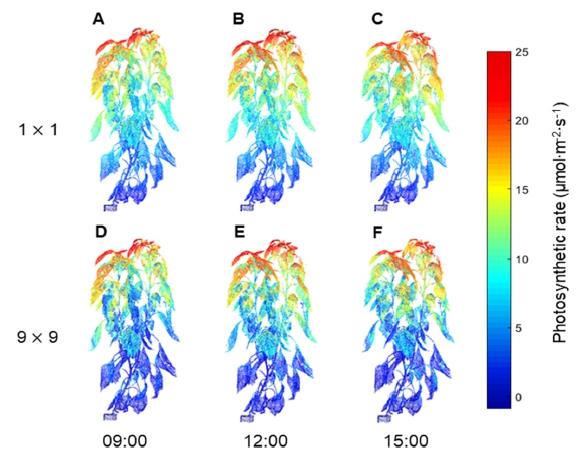
Table 3. Total canopy light interceptions of 3D-scanned paprika models at different isotropic arrangements and times.
 |
직사광이 높은 각도(76.4°)로 작물에 입사되는 정오의 경우, 대부분의 직사광은 작물 상단부에 흡수되고, 하부 엽들은 상위 엽의 차광에 의해 낮은 수광을 보이게 된 다. 또한 태양광이 거의 머리 위에서 수직으로 입사되기 때문에, 작물 상단부의 수광량은 주변 개체의 증감에 크 게 영향을 받지 않는다. 반면 작물 중단부와 하단부의 경우 인접한 개체에 의해 추가적인 차광이 발생하여 수 광량이 크게 감소한다. 이러한 현상은 9×9 배치에서 더 크게 나타났으며, 이는 주변 개체에 의한 상호 가림 현 상이 증가하였기 때문이라고 판단된다(Hilker 등, 2008). 오전 9시와 오후 3시의 캐노피 총 수광량이 정오에 비 해 더 높은 비율로 감소하는 것 역시 태양고도에 의한 영향이라고 볼 수 있다. 오전 9시와 오후 3시의 경우 태 양고도는 각각 43.9°, 42.9°로 정오에 비해 매우 낮아진 다. 낮은 태양고도 하에서는 직사광이 작물에 비스듬하 게 들어오기 때문에, 작물 수광이 상단부에 집중적으로 나타나지 않고, 중단부와 하단부에 비교적 균등히 빛이 분배된다 (Figs. 2A, 2C). 또한 낮은 태양고도 하에서는 인접한 개체에 의해 차단되는 직사광이 많기 때문에 주 변 개체에 의해 캐노피 수광량이 더 많이 감소하게 된다 (Retkute 등, 2008;Li 등, 2014). 본 연구에서는 이론적 관점에서 작물의 배열 방향이 일정한 조건에서 분석하였 지만, 보다 실용적인 결과를 도출하기 위해서는 다양한 방향에 따른 결과의 해석이 필요할 것으로 판단된다.
2. 캐노피 광합성 분포 및 CO2 소모량
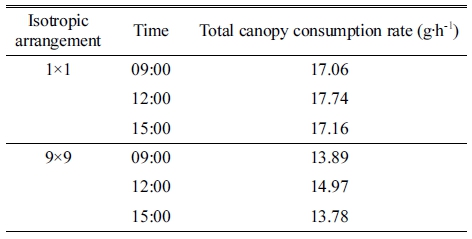
엽 광합성 모델인 직각쌍곡선 모델을 이용하여 광학 시뮬레이션을 통한 파프리카 3차원 스캔 모델의 표면 수광 분포로부터 광합성 속도 분포를 계산할 수 있었다 (Fig. 3). 광합성 속도는 작물 상단부에서 가장 높았고, 하단부로 이르기까지 감소하는 양상을 보였다. 특히 작 물 최하단부 엽은 극히 낮은 광합성 속도를 보였다. 광 합성 분포를 통해 캐노피 CO2 소모량을 계산한 결과 1×1 배치 시 정오에 17.74g·h-1로 가장 높은 값을 보였 고, 오전 9시와 오후 3시에는 각각 17.06, 17.16g·h-1로 비슷한 값을 보였다(Table 4). 9×9 배치 시 캐노피 CO2 소모량은 오전 9시, 정오, 오후 3시에 각각 13.89, 14.97, 13.78g·h-1으로 1 x 1 배치에 비해 각각 81.4%, 84.4%, 80.3%로 감소하였다. 캐노피 CO2 소모량 역시 총 수광량과 비슷하게 정오에 가장 높은 값을 나타내었 다. 반면 정오에서의 작물 상단부 평균 수광량이 약 1,979.3μmol·m-2·s-1로 각각 1,099.2, 1,096.1μmol·m-2·s-1였 던 오전 9시와 오후 3시에 비해 약 1.8배 가량 높았음에 도 불구하고 캐노피 CO2 소모량에는 큰 차이가 나타나지 않았다. 이는 400μmol·mol-1의 CO2 농도에서 엽 광합성 속도가 포화되기 때문일 것으로 생각된다. 직각쌍곡선 모 델을 이용하여 캐노피 상단부 엽의 광합성 속도를 계산 한 결과 400μmol·mol-1의 CO2 농도에서는 광합성 유효 광양자속 밀도 1,200μmol·m-2·s-1에서 23.3μmol·m-2·s-1의 광합성 속도를 보이며, 광합성 유효 광양자속 밀도 2,000μmol·m-2·s-1일 때는 25.6μmol·m-2·s-1의 광합성 속도 를 보였다. 따라서 광도가 약 67% 증가하였음에도 광합 성 속도는 약 9% 정도의 작은 증가폭을 보였다.
Table 4. Total canopy CO2 consumption rates of 3D-scanned paprika models at different isotropic arrangements and times.
 |
작물 수광에는 다양한 변수가 영향을 미친다(De Visser 등, 2014). 작물 개체 내에서는 엽의 분포, 엽각 등의 구조적 형태에 의해 수광 패턴이 변화하며, 인접한 개체에 의한 상호 차광 역시 작물 수광에 영향을 미친 다(Tanaka and Kawano, 1966;Kim 등, 2016). 따라서 작물 표면의 수광을 정확히 측정하는 것에는 큰 어려움 이 있다. 이번 실험에서는 이에 대한 적절한 대안으로 작물 3차원 스캔 모델과 광학 시뮬레이션을 이용한 방 법을 선택하였다. 파프리카의 3차원 스캔 모델을 대상으 로 한 광학 시뮬레이션 결과, 특정 환경 하에서의 작물 표면 수광 분포에 대한 예측이 가능하였으며, 시뮬레이 션 결과로 얻은 수광 분포에 엽 광합성 모델을 적용하 여 광합성 속도 분포 역시 계산할 수 있었다. 본 연구의 분석에서는 가상의 시뮬레이션 환경을 이용하여 온도, 상대습도 등의 환경 변수를 일정하게 고정하였기 때문에 작물이 재배되는 온실 내의 실제 환경을 정확히 반영하 지는 못했다는 한계가 있었으나, 3차원 작물 모델과 광 학 시뮬레이션을 이용하여 작물 수광과 광합성 속도의 공간적 분포를 예측하고 계산해 볼 수 있었다. 따라서 본 연구에서의 예측 방법론은 추후 다양한 환경 하에서 의 작물 수광과 캐노피 광합성의 분석에 대한 새로운 가능성을 제시하였다고 사료된다.
