

서 론
재료 및 방법
1. 실험 환경
2. 실험 작물
3. 데이터 수집
4. Clinostat(클리노스테이트)
5. 데이터 분석 방법
결과 및 고찰
1. 엽장 및 엽폭
2. 엽록소 형광량(OJIP) 분석
3. 광화학적 생리 지표 분석
서 론
우주농업은 지속 가능한 우주 탐사와 정착을 위해 필수적인 연구 분야로 자리잡고 있다. 우주에서 장기간 체류하기 위해서는 안정적인 식량 공급이 필요하며, 이를 위해 중력이 약한 우주환경에서 식물을 재배하는 기술의 확보가 요구된다(NASA, 2018). 현재까지 수행된 연구들은 대부분 국제우주정거장(ISS) 및 지구 내에서도 미세중력환경을 모사한 장치 및 미세중력에서의 식물의 반응을 연구 중에 있다.
지구와 달리 우주 환경에서는 중력, 방사선, 기압 등 여러 물리적 요소가 크게 다르며, 이는 식물의 생장과 생리적 반응에 중요한 영향을 미친다(Zhen, 2015). 중력 변화는 식물의 세포 구조, 신호 전달 과정, 뿌리 및 줄기의 성장 방향뿐만 아니라 광합성 과정에도 영향을 미친다(Kochubey 등, 2007). 특히, 광합성 반응의 효율성은 식물의 생장과 생산성을 결정짓는 중요한 요소이며(Kochubey 등, 2007), 중력 변화가 이에 미치는 영향을 평가하는 것은 우주농업 시스템 구축에 있어 필수적인 연구 분야이다.
국제우주정거장과 지상 기반 미세중력 시뮬레이션 플랫폼(Herranz, 2014)을 활용한 연구들을 통해 식물 생물학적 반응에 대한 이해가 진전되고 있다. 예를 들어, 미세중력이 세포벽 변화(Hoson 등, 2002; Johnson 등, 2015)와 세포 주기(Manzano 등, 2009)와 같은 변화된 표현형의 세포 및 분자적 변화에 미치는 영향이 상세히 규명되었다. 이러한 변화들을 분석하기 위해, 미세중력 환경을 모사할 수 있는 클리노스테이트가 활용되고 있으며, 이를 통해 식물의 중력 적응이 관찰되고 있다. Arabidopsis thaliana를 대상으로 한 클리노스테이트 연구에서는 중력 감지 및 식물 반응과 관련된 유전자(HY5, MIZ1, microRNA MIR163)의 mRNA 발현량 감소가 확인되었다(Wang 등, 2016). Mouhamad 등(2016)은 클리노스테이트 기반 실험을 통해 중력 변화가 단순한 생장 반응을 넘어 아미노산 합성과 같은 대사 과정에도 미친다는 사실을 보고하였다. Hasenstein과 van Loon(2015)은 클리노스테이트와 같은 회전 시스템의 설계 및 기능을 상세히 분석하면서, 이러한 시스템이 미세중력 연구에서 어떻게 활용되는지와 그 한계점을 논의하였다. Nishimura(2023)에 따르면, 클리노스테이트 및 무작위 위치 결정 장치(Random Positioning Machine, RPM)와 같은 미세중력 모사 기술은 생명과학 연구뿐만 아니라 줄기세포 연구에도 활용되며, 중력 변화가 세포 성장 및 분화에 미치는 영향을 연구하는 데 중요한 도구가 되고 있다. 실제 우주에서도 실험을 통해 NASA의 VEGGIE 프로젝트에서는 국제우주정거장(ISS)에서 상추와 밀을 재배하며 미세중력이 식물 생장에 미치는 영향을 연구하였다(NASA, 2019).
미세중력 환경에서는 광합성에 관여하는 PSⅡ의 구조 및 기능이 변형될 수 있으며, 이는 식물의 생장뿐 아니라 광합성 효율 전반에 영향을 줄 수 있다. 엽록소 형광은 이러한 광합성 시스템의 상태를 비파괴적으로 진단할 수 있는 도구로, 특히 는 PSⅡ의 최대 양자효율을 나타내며, 환경 스트레스에 따른 광합성 장치의 민감한 반응을 반영한다. 기존의 연구들에서는 미세중력 조건에서 PSⅡ, PSⅠ, 전체 전자전달(WCE)의 활성이 감소하고, 비율이 지상 조건에 비해 낮아지는 경향을 보였으며, 이는 고광량 환경에서 더욱 뚜렷하게 나타나는 것으로 보고되었다(Stutte 등, 2005).
이에 따라 미세 중력 하에서 엽록소 형광 특성과 광합성 시스템의 기능 변화를 체계적으로 분석할 필요성이 대두된다.
본 연구는 기존 연구와 차별화된 접근으로, 지상에서 클리노스테이트를 활용해 단계별 중력 상쇄 조건을 설정하고, 이를 통해 식물의 생장 및 광합성 반응을 비교 분석하고자 한다.
실험에 사용된 2-D 클리노스테이는 중력 자극에 대한 식물의 생리적 반응, 광합성 생장 및 엽록소 형광 반응을 분석하기 위한 목적으로 활용되었으며, 클리노스테이트는 수평·수직 축 회전을 통해 일정 방향의 중력 자극을 상쇄하며, 중력 벡터를 시간적으로 평균화시켜 미세중력 환경을 모사한여, 중력이 식물 생장과 광합성 기능에 미치는 영향을 정량적으로 평가할 수 있는 효과적인 실험 도구로 사용된다.
상추는 상대적으로 빠른 생장속도와 간단한 생육 관리가 가능하여 우주농업 연구에서 모델 식물로 선호된다. 또한, 상추는 광합성 효율을 평가하는데 적합한 종으로 엽록소 형광 분석을 통해 중력 변화에 따른 생리적 반응을 명확히 확인할 수 있다.
따라서, 본 연구는 상추를 대상으로 클리노스테이트 회전축으로부터의 거리 차이에 따른 생리적 반응을 비교 분석함으로써, 중력 변화가 생장 및 엽록소 형광 특성에 미치는 영향을 평가하고자 한다. 이를 통해 중력 변화가 식물의 생장과 광합성에 미치는 영향을 정량적으로 평가하고, 향후 우주에서의 식물 재배 시스템 최적화에 필요한 기초 데이터를 제공하고자 한다.
재료 및 방법
1. 실험 환경
본 연구는 경상국립대학교에 설치된 반폐쇄형 식물공장에 클리노스테이트를 설치하여 수행되었다(Fig. 1). 식물공장의 전체 규모는 3m, 세로 6m, 높이 2.8m이며, 내부는 좌우로 각각 9개의 배드로 구성되어 총 18개의 재배 공간으로 분류된다. 각 배드의 크기는 가로 1,236mm, 세로 616mm, 높이 700mm로 구성되어 있으며, 본 실험은 중앙에 위치한 배드에서 수행되었다. 내부 환경은 자동 제어 시스템을 통해 안정적으로 유지되었으며, 온도는 주간20°C, 야간 18°C로 설정하였고, 상대습도는 60 ± 5% 수준으로 유지되었다. 광원은 LED를 사용하였으며, 450nm의 청색광과 660nm의 적색광을 1 : 2의 비율을 구성하였다. 광주기는 상추의 광양 생육 조건으로 12시간 명기와 12시간 암기로 구성된 24시간 광주기를 적용하였으며, 명기는 오전 06시부터 오후 18시까지로 설정하였다. 이는 상추의 생리적 일주기 리듬에 부합하며, 광합성 및 생육에 안정적인 조건을 제공하기 위함이다. Park 등(2012)은 광주기 12/12h 조건에서 상추의 엽록소 형광값이 가장 높게 나타났다고 보고하였고, 이에 따라 광주기 조건을 설정하였다.

2. 실험 작물
오향적치마 상추는 국내에서 재배되는 적엽계 상추 품종으로, 생장 속도가 빠르고 광합성 활성도가 높아 실험에 적합한 작물로 판단되어 선정하였다.
파종은 2024년 9월 24일에 실시하였으며, 생육이 일정 수준 도달한 이후인 2024년 10월 5일부터 9일간 본 실험을 수행하였다.
양액이나 양분은 별도로 제공하지 않았으며, 물은 하루에 한 번 아침 9시에 10mL씩 제공하였고, 각 개체의 무게는 포트를 포함하여 250g ± 5%로 설정하였다. 각 그룹은 5개의 포트를 클리노스테이트 회전축의 중심점을 기준으로 각기 다른 거리에 배치하여 생장과 엽록소 형광 특성을 비교 분석하였다.
3. 데이터 수집
본 연구에서는 클리노스테이트 중력 상쇄 조건에 따른 식물의 생장 반응 및 생리적 활성을 평가하기 위하여 엽장, 엽폭, 엽록소 형광을 주요 조사 항목으로 선정했다. 이 지표들은 식물의 형태적 성장뿐 아니라 광합성에 대한 생리적 반응을 반영하는 중요한 특성으로 중력 변화에 따른 반응성을 민감하게 나타낼 수 있기 때문이다(Kawamaoto와 Morita, 2022).
본 실험에서는 각 처리구 식물체의 가장 발달한 본엽과 아차엽을 측정하였다. 엽폭은 동일한 잎의 가장 넓은 지점을 기준으로 엽맥에 수직인 방향의 폭을 측정하였다. 각 포트당 일 1회 동일한 잎 위치에서의 엽장과 엽폭을 측정하여 그룹 간의 생장 차이를 정량적으로 비교하였다. 모든 측정은 디지털 버니어 캘리퍼스를 이용하였다.
엽록소 형광은 광합성율과 PSⅡ의 기능 상태를 반영하는 지표로, 스트레스 민감성 평가 및 생리적 활력 측정에 유용하다(Maxwell과 Johnson, 2000). 본 연구에서는 휴지기 형광(F₀), 최대 형광(Fₘ), 유도 형광 곡선, JIP test 지표 등 다양한 형광 파라미터를 활용하여 중력 변화에 따른 식물의 생리적 반응을 정량적으로 분석하였다. 엽록소 형광 측정기 OS-30p+ (ADCBioScientifil Ltd, UK)를 이용하여 엽록소 형광 측정기의 측정 매뉴얼에 따라 암적응이 30분 이상 유도한 후 오후 19시 가량에 실험 기간동안 매일 동일한 시각에 측정하였다.
4. Clinostat(클리노스테이트)
본 실험은 RB_35GM 21TYPE (12V)를 회전 모터로 사용하였고 회전속도는 7 RPM으로 설정하였다. 식물 포트를 고정하기 위해 유리 원판을 사용하였다.
클리노스테이트를 이용한 실험에서는 중력 상쇄 효과를 적절히 평가하기 위해 1g 대조군을 신중하게 설정하는 것이 중요하다(Kamal 등, 2015). 이를 통해 클리노스테이트 실험에서 대조군 설정의 중요성을 반영하였으며, 기존 연구에서 제안된 방식과 유사한 방법을 적용하였다.
본 실험에서는 총 4개의 그룹으로 구분하여 실험을 진행하였다. 대조군으로 클리노스테이트(Clinostat)에 부착하지 않은 그룹과 클리노스테이트 회전축의 중심(0cm), 중심으로부터 6cm, 12cm 떨어진 위치에 배치된 그룹을 설정하였다
여기서,
a : 원심 가속도
ω : 각속도 (rad∕s)
r : 회전축 중심으로 부터의 반지름 거리 (m)
g : 중력 가속도 (9.81 m/s²)
클리노스테이트의 회전 속도를 7RPM으로 설정하였을 때, 각속도는 다음과 같이 계산된다.
본 실험에서 측정한 반경은 클리노스테이트 중심(0cm), 중심에서 6cm, 12cm 떨어진 지점이며 이를 미터 단위로 변환하여 적용하였을 때, 각 지점에서 원심 가속도는 다음과 같이 계산된다.
실험 조건에 따라 각 군은 다음과 같이 구분하였다. 회전이 없는 대조군(control group)은 acon, 클리노스테이트 중심부(반경 0.001m)는 acen, 반경 6cm 위치는 a6, 반경 12cm 위치는 a12로 정의하였으며, 이후 모든 분석에서 동일한 표기법을 적용하였다.
위의 결과를 클리노스테이트 회전 반경과 중력 상쇄 효과를 나타내는 Hasenstein과 Van Loon(2015)의 중력 상쇄 그래프와 비교한 결과, 위에서 계산한 값과 그래프의 위치해 있는 값이 유사하게 나타났다.
중심 (0.001m, 최소값 가정) :
6cm(0.06m) :
12cm(0.12m) :
5. 데이터 분석 방법
9일 동안 일 1회 데이터를 수집하여 각 그룹당 3 포트를 사용하여 엽장, 엽폭 및 엽록소 형광 데이터를 분석하였다. 각 그룹별 엽장과 엽폭의 변화를 비교하여 클리노스테이트와 중심으로부터 위치한 거리가 식물의 형태적 성장에 미치는 영향을 평가하였으며 이를 위해 일자별 평균값을 도출하고 시간에 따른 변화를 그래프로 시각화하여 비교하였으며, 광화학적 생리 지표 분석으로는 반응 중심 활성도, 단면적당 광합성 활성 변화를 포함한 JIP-Test 지표를 사용하여 분석하였다.
수집된 데이터는 그룹 간 차이를 시각적으로 비교하기 위해 그래프로 표현하였으며 엽장 및 엽폭 변화, 엽록소 형광 반응 및 JIP - Test 변수 간 변화를 비교 분석하였다. 특히, 클리노스테이트 회전축과의 거리 차이가 식물의 생장 및 광합성 시스템의 적응에 미치는 영향을 정량적으로 평가하였다. 엽장 및 엽폭 변화는 일자별 평균값 및 증가율을 비교하여 식물의 형태적 성장이 클리노스테이트 환경에서 어떻게 적응하는지를 분석하였으며, 엽록소 형광 광계 기능에 미치는 영향을 확인하기 위해, 시간 경과에 따른 광합성 반응 변화를 확인하였고, 또한, 엽장과 엽폭 데이터의 통계분석은 SPSS(IBM Corp., Armonk, NY, USA)를 이용하여 일원배치분산분석(One-way ANOVA)를 실시하였으며, 유의성이 나타난 경우 Duncan의 다중 범위 검정(Duncan’s multiple range test)를 이용하여 사후 검증을 통해 클리노스테이트 회전축과의 거리에 따른 중력 차이가 식물의 생리적 반응에 미치는 영향을 정량적으로 분석하였다.
Table 1은 실험 중 측정한 엽록소 형광 데이터 중 분석에 활용된 주요 지표들을 요약한 것이다.
Table 1.
Formulas and glossary of terms used by the JIP-test for the analysis of the fluorescence transient OJIP.
Fig. 2은 발아된 상추를 토양 용기에 심은 후 클리노스테이트에 설치한 상태이다.

결과 및 고찰
1. 엽장 및 엽폭
Fig. 2는 클리노스테이트에 부착되어 회전하고 있는 상추의 모습이다.
엽장 같은 경우 Days after treament(DAT) 1부터 DAT 9까지 모든 처리구에서 유의적인 차이가 나타나지 않았다(Fig. 3). 실험 기간 동안 클리노스테이트의 중력 감소 효과에 따른 엽장의 차이는 크지 않았으며, 모든 처리구가 유사한 성장 패턴을 보였고, 처리에 따른 엽장의 변화가 뚜렷하게 나타나지 않았음을 보여준다.
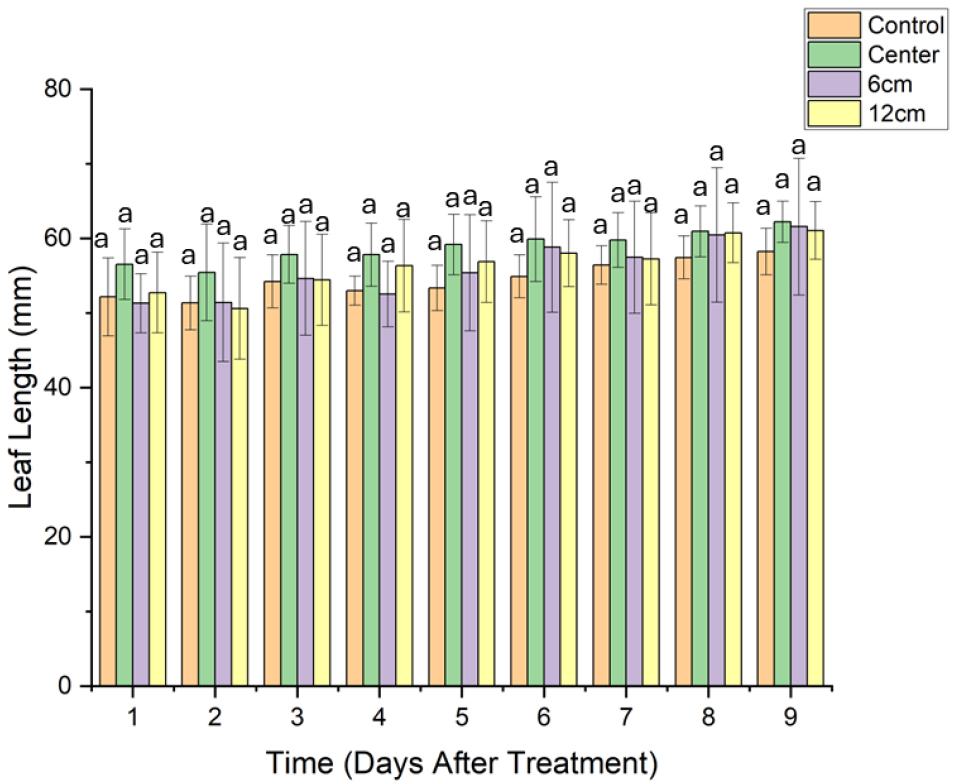
엽장과는 다르게 엽폭에서는 일부 처리구간에서 유의미한 차이가 확인되었다. a5, a12 처리구에서 다른 처리구과 유의한 차이를 보였다(Fig. 4). 이는 a6, a12 위치에서 자란 식물의 엽폭이 다른 처리구보다 작을 가능성이 높음을 의미한다.
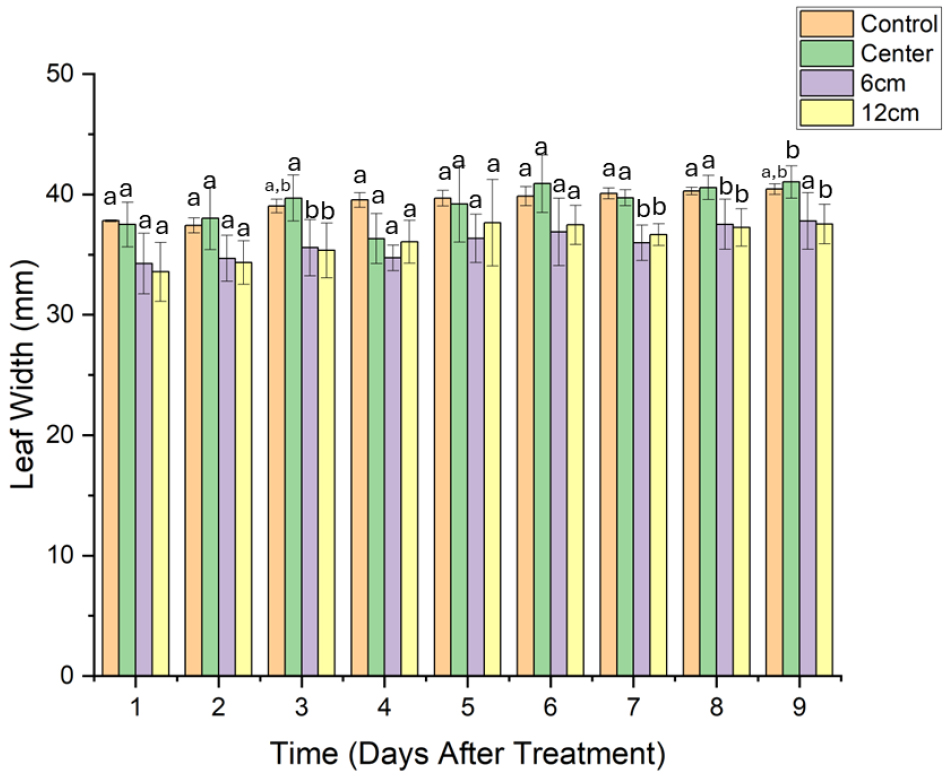
DAT 3에서는 a6 및 a12 처리구가 acon 및 acen 대비 유의하게 낮게 나타났다. 이는 회전축에서 멀어질수록 작용하는 원심력의 영향으로 식물체가 중력 방향을 인식하는 기작에 혼란을 겪었거나, 물·양분 분포의 균형이 깨져 잎의 수평 확장이 저해되었을 가능성이 보인다.
DAT 1부터 DAT 9까지의 데이터를 확인한 결과, a6 및 a12 그룹은 다른 그룹에 비해 실험 후반부에도 낮은 경향을 보였다.
또한, 모든 처리구에서 유의적인 차이가 나타나지 않은 DAT 2, DAT 4-DAT 6는 실험 초반과 일부 시점에서 클리노스테이트의 환경이 엽폭에 미치는 영향이 뚜렷하지 않다는 것을 보여줬다. 그러나 후기 반응 단계(DAT 7-DAT 9)에서 a6와 a12 처리구가 acon 및 acen 처리구와 비교하여 유의하게 낮을 값을 보였다는 점에서 시간이 지남에 따라 클리노스테이트의 위치에 따른 차이가 발생할 수 있음을 보여준다.
2. 엽록소 형광량(OJIP) 분석
Table 1은 엽록소 형광량 분석에 사용된 변수들을 보여주며 를 사용하였다.
Fig. 5는 1일차부터 9일차까지 기록된 평균 엽록소 형광 OJIP 곡선을 나타낸 것으로 x축은 로그 스케일(Time, Day)로 표시되었으며, y축은 형광 강도(a.u.)를 나타낸다. 이러한 곡선은 PSⅡ의 활성과 효율성, 그리고 미세중력 환경 하에서 전반적인 광합성 성능을 반영한다(Jagtap 등, 2024).
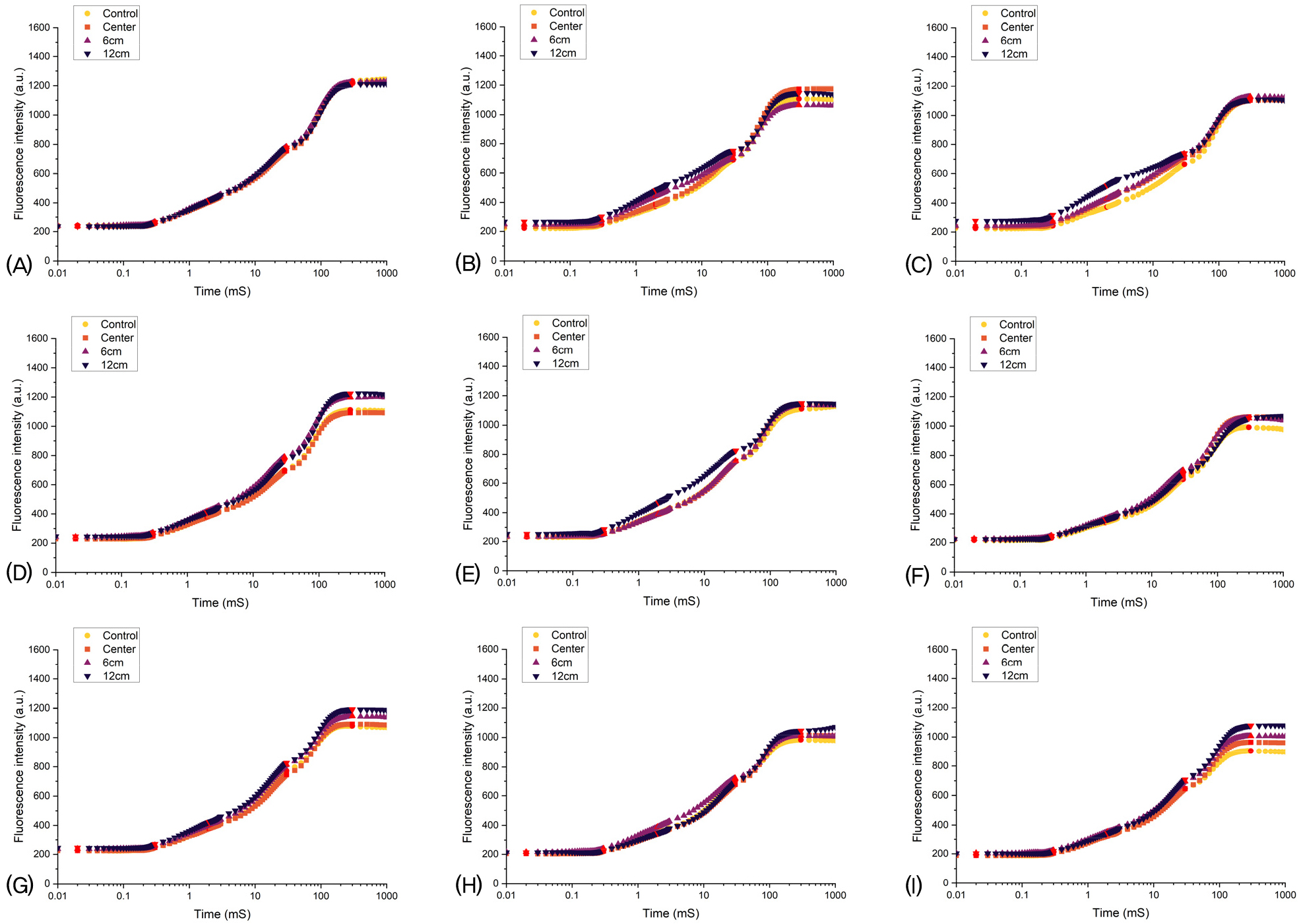
Day 1의 경우, 모든 처리구에서 초기(0-0.1초) 형광 값이 급격히 상승하는 것이 확인되었다. 빛을 받은 PSII에서 가 전자를 받아 광합성 초기 반응이 활성화되었음을 의미한다. 그러나, 시간이 지남에 따라 그룹별 차이가 나타났다. acon과 acen그룹은 안정적인 형광 패턴을 보이며, 약 1.5초 이후 형광 값이 평형을 이루었다. 반면, a6와 a12 그룹에서 형광 값이 높게 유지된 것은, 중력 감지 및 세포 내 응력 반응으로 인해 틸라코이드 막 내 전자전달 복합체의 배열이 불안정해졌거나, 스트레스 반응에 의해 PSⅡ의 안정성이 저하되었기 때문으로 보인다. 이는 정상적인 경로가 차단되면서 과잉 에너지가 형광 형태로 방출되었음을 시사하며 클리노스테이트 외곽부 식물에서의 광에너지 처리 능력 감소를 반영한다. J 단계(2ms)는 Day 3일차부터 a6 및 a12 그룹이 acon보다 높은 형광 값을 보였으며, 가 정상적으로 전자를 전달하지 못하고 축적되었을 가능성을 보여준다.
Maximal recorded fluorescence(P 단계)에서는 acon이 가장 낮고, a6가 가장 높은 형광 값을 보였으며, 이는 과도한 에너지가 형광으로 방출될 가능성이 있음을 보여준다.
9일 동안의 형광 데이터를 분석한 결과, acon과 acen 그룹은 안정적인 형광 패턴을 유지하였으나, a6와 a12 그룹은 변동성이 크게 나타났다. Day 5에서는 a6 및 a12 그룹에서 형광 값이 급격히 증가하였으며, 이는 가 정상적으로 전자를 전달하지 못하고 형광 방출이 증가했을 가능성을 의미한다. 이는 중력 변화가 광합성 및 전자전달계뿐만 아니라 세포 신호 전달 과정에도 영향을 미칠 가능성을 보여준다.
OJIP 곡선 분석 결과, a6 및 a12 그룹에서 P 단계에서의 형광값이 acon보다 높게 나타났으며, 이는 광합성 전자전달 과정의 효율이 저하되었을 가능성을 시사한다. 이는 Kordyum과 Adamchuk(1997)의 연구 결과와 유사한 패턴을 보이며, 클리노스테이트 회전이 틸라코이드 막의 상태 변화 및 광합성 전자전달계 기능에 영향을 미칠 수 있음을 보여준다.
3. 광화학적 생리 지표 분석
광화학적 생리 지표 분석을 위해 사용된 변수들로는 Table 1에 표시된 변수들을 사용하였다.
Fig. 6은 서로 다른 광합성 지표를 나타내며, 각 점은 해당 일자의 각 그룹의 평균값을 의미한다. 이러한 지표들은 다양한 중력 조건 하에서 PSⅡ의 활성과 효율을 종합적으로 반영한다.
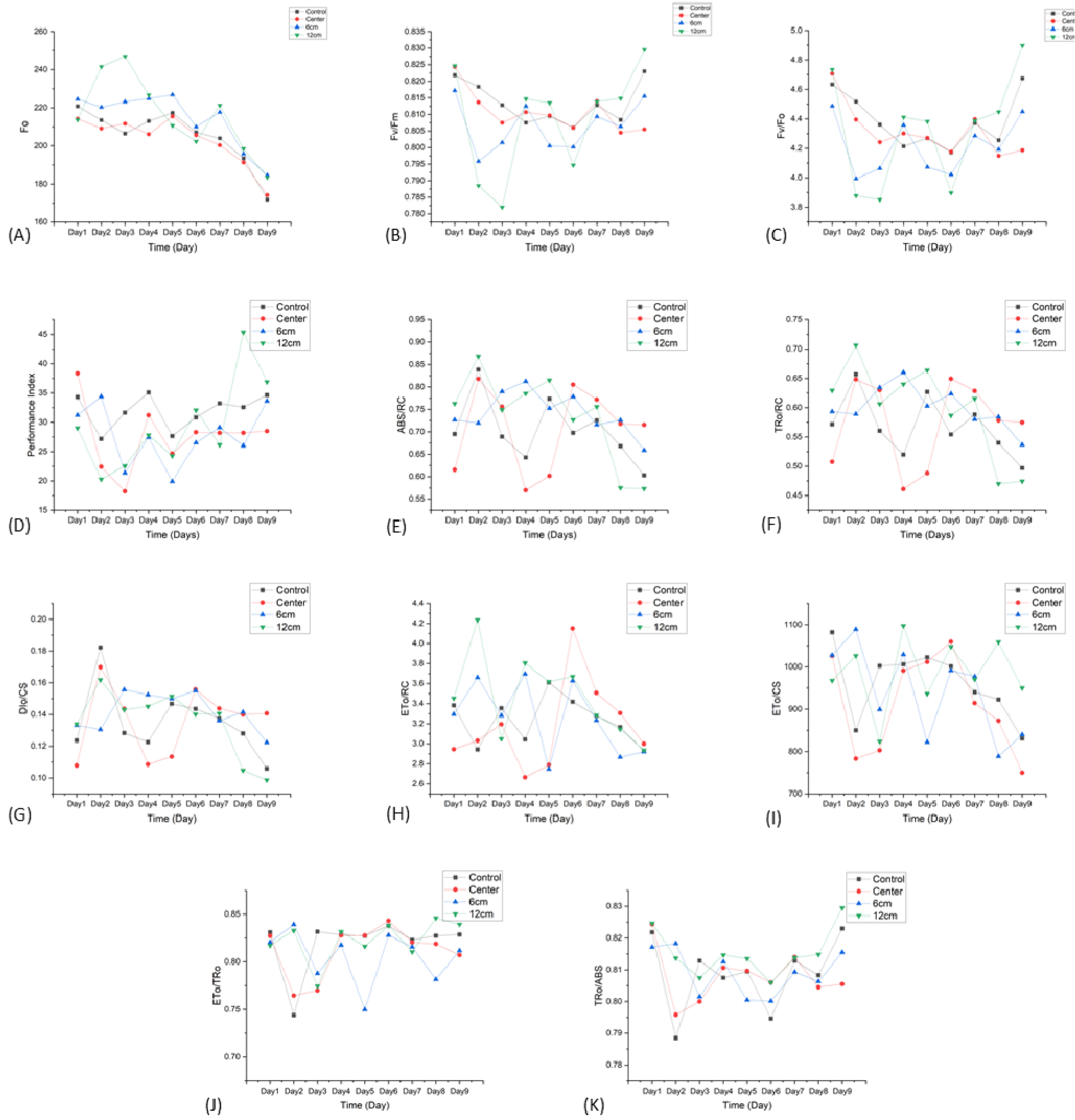
본 연구에서는 클리노스테이트 회전 환경에서의 위치 변화가 식물의 광화학 반응에 미치는 영향을 분석하기 위해 acon그룹을 기준으로 acen, a6, a12 처리구 간의 엽록소 형광 매개변수를 비교하였다.
은 acon과 acen 그룹에서 전반적으로 안정적인 수준을 유지하였으나, a6 및 a12 그룹은 Day 2-4 사이에서 값이 일시적으로 상승한 후, 후반부로 갈수록 급격한 하락을 나타냈다. 이는 Murchie와 Lawson(2013)이 제시한 바와 같이, 의 상승이 PSⅡ 반응 중심의 일시적 폐쇄 혹은 에너지 과부하에 따른 스트레스 반응을 의미하며, 이후의 감소는 광계 손상 또는 안테나 단백질의 손실로 인한 PSⅡ 기능 저하를 반영하는 것으로 해석된다.
은 acon 그룹에서 안정적인 값을 유지하였고, PSⅡ가 정상적인 상태를 유지하고 있음을 의미한다. acon 그룹은 Day 2-3에서 일시적으로 하락한 후 회복 정체를 보였으며, 후반부에는 일정 수준으로 회복되었다. 반면 a6 및 a12 Day 2-3부터 의 값이 급격히 저하되었고, 회복되는데 시간이 소요되었으며, 실험 종료 시점까지도 acon 수준에는 도달하지 못하였다. 이는 회전축으로부터 멀수록 PSⅡ의 스트레스 민감도가 높았고, 회복력은 낮았음을 보여준다.
값 역시 acon 그룹에서 안정적으로 유지되었으며. acen 그룹은 Day 2-4 기간동안 일시적으로 값이 낮아졌으나 이후 회복세를 보였다. 반면, a6 및 a12그룹에서는 의 값이 급격히 감소한 시점이 관찰되었고, 특히 a12 그룹은 실험 후반에서 급격한 반등이 보였다. 회전 조건이 PSⅡ 활성에 큰 영향을 미쳤으며, 특히 회전 축에서 먼 위치에 있는 식물은 광합성 반응 중심의 활성도가 현저히 저하되었다가 뒤늦게 회복되는 패턴을 보였다.
이와 유사하게, Makhtoum 등(2023)의 연구에서도 가뭄 및 염분 스트레스 조건에서 와 값이 유의미하게 감소하였으며, PSⅡ의 기능 저하와 관련된 것으로 보고되었다.
PI는 그룹 간 차이가 가장 뚜렷하게 나타난 지표 중 하나였다. a6과 a12 그룹은 Day 1부터 값이 급격히 감소하였고, 이후에도 낮은 수준을 지속하여 회복되지 않았다. 이는 클리노스테이트 회전축으로부터 멀어질수록 중력 벡터 변화가 커지고, 이에 따라 PSⅡ의 반응 중심이 불안정해져 전자전달 효율이 저하되었음을 의미한다. 화전 반경이 큰 구간은 중력 방향의 지속적인 교란으로 인해 생리적 스트레스가 누적되어 광화학 시스템의 회벅이 지연된 것으로 해석된다.
반면 acen 그룹은 Day 2-3에서 일시적인 PI 하락이 있었으나, 실험 중반 이후 acon과 유사한 수준으로 회복되었으며 회전 중심 위치의 식물이 환경 변화에 일정 시간 내 적응할 수 있음을 보여준다.
반응 중심 단위당 에너지 흡수 및 전달 능력을 나타내는 , 는 a6과 a12 그룹에서 일관된 감소가 확인되었다. 이는 반응 중심이 광 에너지를 수용하거나 전자 전달에 활용하는데 있어 효율이 저하되었음을 시사한다. acen그룹은 초기에는 약간의 변동을 보였으나 빠르게 안정화되었으며, 회전 중심 위치가 광 에너지 처리 능력 유지에 긍정적으로 작용했음을 알 수 있다.
이와 같은 반응 중심 단위당 에너지 수용 및 전달 능려그이 저하는 PSⅡ의 광화학 반응 효율 저하로 이어지며, 이는 광합성 에너지 변환 과정의 전체 효율에 부정적인 영향을 미친다(Kalaji and Guo, 2008).
반면 단위 면적당 소실 에너지를 나타내는 는 a6과 a12그룹에서 상대적으로 높은 값을 보였고, 특히 a12 그룹은 실험 후반까지 소실 에너지가 증가하는 경향이 두드러졌다. 이는 회전으로 인한 물리적 스트레스가 광 에너지의 효율적 활용을 저해하고 손실을 증가시켰다는 결과로 해석된다. acen 그룹 역시 초기에는 값이 다소 상승했으나 이후 안정화되었으며 생리적 회복 양상을 보았을 때 적응 가능성이 크다고 볼 수 있다.
전자 전달과 관련된 역시 a6 및 a12 그룹에서 전체적으로 낮은 값을 나타냈으며. 특히 a12 그룹에서는 광합성계 전자 흐름이 상당이 억제된 상태로 유지되었다.
이는 미세중력 조건에서 PSⅡ에서 시작된 전자가 정상적인 전달 경로를 따라 최종 수용체까지 도달하지 못하고 중간 단계에서 정체되었음을 의미한다.이러한 현상은 중력 변화에 따라 틸라코이드 막의 구조적 안정성이 약화되거나 전자전달 복합체 간의 배열이 무너졌을 가능성과 밀접하게 관련된다. 회전축에서 멀어질수록 중력 벡터의 변화가 더 커지기 때문에 전자전달계의 기능적 안정성이 더욱 저하되고 PSⅡ단위의 활성이 감소하여 광합성 전체 효율이 떨어진 것으로 해석된다.
광 에너지 전환 효율을 나타내는 지표인 은 a6 및 a12 그룹에서는 두 지표 모두 전반적으로 낮은 값을 유지하여 흡수된 에너지가 전자 전달로 연결되지 못하고 손실되는 비율이 높았음을 의미한다. 특히 비율은 a12 그룹에서 가장 낮았으며, 이는 에너지 변환 체계의 효율성 자체가 크게 저하되었음을 보여준다. acen 그룹은 초기 일시적 하락 후 회복되는 경향을 보였으며, 전반적으로 적응 가능성이 높게 나타났다.
결과적으로, 클리노스테이트에서의 그룹 별 차이는 회전축에서 멀어질수록 광화학적 생리 지표의 변화폭이 커지고, PSⅡ의 기능 저하가 뚜렷하게 나타났다. 반면 회전 중심에 위치한 식물은 초기 스트레스를 경험하였으나 빠르게 안정화되어 대조군과 유사한 생리 상태로 회복되었으며, 이는 광화학적 생리 반응의 클리노스테이트의 위치 의존성을 보여준다. 이러한 결과는 중력 변화 환경에서 식물의 광합성 반응을 해석하는데 있어 회전 조건과 위치의 중요성을 보여주며, 향후 중력 유사 환경 연구의 설계에 있어 활용 가능할 것으로 사료되었다.
