

Introduction
Materials and Methods
1. Seed characteristics
2. Seed imbibition
3. Temperature and light treatments
4. Germination parameters
5. Statistical analysis
Results and Discussion
1. Seed characteristics and imbibition
2. Germination parameters
Conclusion
Introduction
Coriandrum sativum L., commonly called as coriander or cilantro, is an annual herbaceous species in the Apiaceae. As one of the oldest aromatic herbs, it has a 3,000-year history of use in culinary, medicinal, and cosmetic applications (Kumar et al. 2022). Coriander is characterized by its strong and distinctive aroma, which is largely attributed to aldehydes and volatile metabolites such as linalool, -pinene, borneol, camphor, terpinene, phthalides (Naeem et al. 2022). In the food industry, its leaves and stems are consumed fresh or as ingredients in cooked dishes, whereas its seeds are primarily used as a spice to enhance food flavor and aroma. In addition, coriander contains diverse bioactive metabolites, notably phenolic acids (gallic, caffeic, and chlorogenic acid) and flavonoids including flavone, luteolin and kaempferol (Gantait et al. 2022; Kumar et al. 2022; Naeem et al. 2022). It is also rich in vitamins, minerals, and dietary fiber, containing calcium (0.14%), phosphorus (0.06%), iron (0.01%), vitamin B2 (60 mg·100 g-1), vitamin C (135 mg·100 g-1), and vitamin A (10,460 IU·100 g-1) (Nadeem et al., 2013). Owing to these nutritional and phytochemical properties, coriander plant has been reported to provide diverse health-promoting effects, including antioxidant, antidiabetic, antibacterial, antifungal, anti-inflammatory, and cytotoxic activities, and has traditionally been used to treat ailments such as stomachache, cough, and diarrhea (Khan and Khatoon 2008; Mahleyuddin et al. 2021).
Harvest season and yield of coriander differ markedly among producing countries. Notably, November and December fall within the off-season in several production regions, and this seasonal gap often results in unstable prices and reduced uniformity in the quality of imported coriander (Tridge 2026). Against this background, domestic coriander production in Republic of Korea has attracted increasing interest. The cultivation area expanded from approximately 34.5 ha (2017) to 54.2 ha (2022), and market prices increased steadily from 9,604 to 11,648, 14,128, and 16,774 KRW·kg-1 in 2019, 2020, 2021, and 2022, respectively (Gi et al. 2024). Therefore, coriander has recently attracted attention as a high-value crop for controlled environment plant production systems, including vertical farms. Consistent with this trend, recent studies worldwide have reported that hydroponic cultivation under controlled environments is an effective and intensive approach for the large-scale production of coriander (Nguyen et al., 2020; Gao et al., 2022; Gi et al., 2024).
Despite the high value of coriander, poor and uneven seed germination remains a major constraint for its large-scale production, largely due to limited water permeability imposed by the hard seed coat and the presence of coumarin, a germination-inhibiting compound in the seeds (Arora et al. 2021; Palani-Vasantha et al. 2024). Palani-Vasantha et al. (2024) reported that coriander seed germination was improved by water imbibition leading to softening the seed coat or leaching the germination inhibitor compounds in seeds. However, coriander seeds are very sensitive to temperature during storage and germination, and under conditions where the temperature is not uniformly controlled, germination becomes uneven and the average germination time becomes longer (Lee et al. 2015). To overcome this constraint, it is essential to understand the physiological mechanisms underlying seed germination.
Seed germination is primarily regulated by environmental factors such as temperature and light. Temperature is a key factor for enzyme activation, respiration, and embryo elongation, and each species has an optimal temperature range for stable germination, which determines germination rate and growth speed (Haj et al. 2022). Most species exhibit high germination rates within this range, whereas germination declines sharply outside it (Cochrane 2020). Seed responses to light vary by seed type and are generally classified as positively photoblastic, negatively photoblastic, or non- photoblastic (Liu et al. 2023). Light promotes and accelerates germination in positively photoblastic seeds, whereas it inhibits germination in negatively photoblastic seeds, for which darkness is more favorable (Shinomura et al. 1994; Wang et al. 2021). Thus, unfavorable temperature or light conditions can lead to delay germination and reduced seedling uniformity, ultimately impairing subsequent growth (Carvalho et al. 2020; Luo et al. 2022).
Previous studies on coriander seeds have mainly examined priming treatments to improve germination (Debbarma et al. 2018; Talat et al. 2019), morphological characteristics and germination behavior during storage (Lee et al. 2015), the effects of different sowing depths on germination (Modhavadia et al. 2025), and the viability and vigor of naturally aged seeds (Kumar et al. 2015). Nevertheless, there have been no systematic rereport, to the best of our knowledge, on the interactive influence of temperature and light on coriander seed germination in the context of large-scale vertical farming. Therefore, this study aimed to determine the appropriate temperature and light conditions for enhancing the germination rate and uniformity of coriander seeds under vertical farming systems.
Materials and Methods
1. Seed characteristics
Prior to the germination test, the one-thousand-seed weight (TSW), length, width, embryo-to-seed ratio (E:S ratio), and vigor of coriander (Coriandrum sativum L.) seed (Danong Co., Ltd., Dongducheon, Republic of Korea) were evaluated. The labeled germination rate provided by the seed company was 60%. TSW was determined by measuring the weight of 100 seeds with an electronic balance (SI-234; Denver Instrument, Bohemia, NY, USA) and calculating the corresponding weight for 1,000 seeds. Seed length and width were measured in 10 seeds using a stereomicroscope (SZ61; Olympus, Tokyo, Japan) coupled with a complementary metal-oxide-semiconductor camera (eXcope F-630; Dixi Sci., Daejeon, Republic of Korea). For evaluation of the E:S ratio, ten seeds were cross-sectioned using a double-edged razor blade, and measurements were taken using a stereomicroscope. Seed viability was determined according to the staining pattern using a tetrazolium test and expressed as a percentage. Fifty seeds were first imbibed in distilled water for 24 h under dark conditions at 20°C. After imbibition, the seeds were cut in half and transferred to a 1% tetrazolium solution (triphenyl tetrazolium chloride; Sigma- Aldrich, St. Louis, MO, USA). The prepared seeds were then placed in Petri dishes and incubated in darkness at 30°C for an additional 24 h.
2. Seed imbibition
The initial moisture content and water absorption capacity of coriander seeds were evaluated by first measuring the fresh weight of 50 seeds cut in cross section. The seeds were then placed in distilled water and maintained under dark condition at 4℃. At 0.5, 1, 1.5, 2, 2.5, 3, 4, 5, 6, 8, and 12 h after the start of imbibition, excess surface moisture was removed carefully with tissue paper, and the weight of the seeds was recorded with an electronic balance.
3. Temperature and light treatments
After soaking in distilled water at room temperature for 24 h, the seeds were surface-sterilized in 1.4% NaClO for 15 min, and then rinsed with running distilled water for about 10 min. Fifty sterilized seeds were placed on two sheets of filter paper in a petri dish (90 × 15 mm; SPL Life Sciences Co., Ltd., Pocheon, Republic of Korea), with four replicates per treatment.
Seeds were incubated for 10 days under five temperature (10, 15, 20, 25, and 30℃) and two light treatments (light and dark). Temperature was controlled using a plant growth chamber (LBI-250M; ESSTELL Co., Ltd., Namyangju, Republic of Korea). For the light treatment, the chamber was maintained under continuous light (24 h photoperiod) with a photosynthetic photon flux density (PPFD) of 60 ± 10 µmol·m-2·s-1 and the petri dishes were covered with lids. For the dark conditions, the lidded petri dishes were additionally wrapped with aluminum foil. Moisture was maintained by adding 1.5 mL of distilled water to each petri dish every two days. Germination parameters were evaluated in a dark room under dim green light generated by a flashlight covered with green paper tape to minimize light-induced stimulation.
4. Germination parameters
Germination was assessed by radicle emergence and recorded daily for 10 days. Based on these observation, germination rate (GR), mean germination time (MGT), time required for 50% germination (T50), and promptness index (PI) were calculated using the following formulas below (Seo et al. 2012; Lee et al. 2015).
ni: number of seeds germinated on day i, S: total number of seeds
ni: number of seeds germinated on day i, ti: the i-th day on which germination was observed,
N: total number of germinated seeds
N: total number of germinated seeds,
Ni: cumulative number of seeds germinated up to just before 50% of N,
Nj: cumulative number of seeds germinated immediately after reaching 50% of N,
Ti: time required to reach Ni,
Tj: time required to reach Nj
T: total number of observation days,
ti: the i-th day on which germination was observed,
ni: number of seeds germinated on day i
5. Statistical analysis
Data are expressed as mean ± standard error from four replicates for each treatment. Statistical analyses were performed using Duncan’s multiple range test in SAS (Statistical Analysis System, version 9.4; SAS Institute Inc., Cary, NC, USA), and significance was accepted at P < 0.05, 0.01, and 0.001. Differences in PI between light and dark conditions were evaluated by a t-test. All figures were prepared using SigmaPlot (version 12; Systat Software Inc., San Jose, CA, USA).
Results and Discussion
1. Seed characteristics and imbibition
Coriander fruits were oval in shape and ranged in color from light to dark brown (Fig. S1). Each fruit contained two seeds arranged on opposite sides, and the embryo is small, slender, and located at the nucleus end of the seed. TSW was 9.46 ± 0.07 g, and the seed length and width were 3.81 ± 0.01 mm and 3.17 ± 0.01 mm, respectively (Table 1). E:S ratio was 0.36 ± 0.01 and seed viability by tetrazolium test was recorded as 99.0 ± 0.01% (Table 1 and Fig. S1).
Table 1
Thousand-seed weight (TSW), size (length × width), embryo to seed ratio (E:S ratio), and viability of the coriander seed used in this study
| TSW (g) | Size (mm) | E:S ratio | Viability (%) |
| 9.46 ± 0.07 | 3.81 × 3.17 ± 0.01 × 0.01z | 0.36 ± 0.01 | 99.0 ± 0.01 |
Previous studies by Sharanagat and Goswami (2014) and Coşkuner and Karababa (2007) showed that the physical characteristics of dried coriander seeds varied depending on seed moisture content. Specifically, they reported seed lengths of approximately 5.06 mm and 4.61-4.74 mm, respectively, and seed widths of approximately 3.27 mm and 3.67-3.93 mm, respectively. TSW ranged from 8.72 to 9.71 g in Sharanagat and Goswami (2014) and from 8.89 to 9.826 g in Coşkuner and Karababa (2007). In the present study, the coriander seeds had a length of 3.81 ± 0.01 mm, a width of 3.17 ± 0.01 mm, and a TSW of 9.46 ± 0.07 g. Although direct comparison of seed dimensions is limited by differences in the reported parameters, the comparable TSW suggests that the seeds used in this study were not markedly different from those used in previous studies. Therefore, extreme variation in seed physical traits was unlikely to have substantially biased the germination results.
The coriander seed imbibition curve reached saturation after about 4 h, with a weight of 1.0447 g (Fig. 1). Seed imbibition is considered critical pretreatment factors for promoting uniform and successful germination (Woodstock 1988). In this study, coriander seeds reached a saturated water uptake state after 4 h of imbibition, suggesting that at least about 4 h of imbibition is required to secure sufficient water for germination initiation. Accordingly, under the present experimental conditions, initial water uptake itself does not appear to be the sole factor limiting germination in coriander seeds. Coriander fruits are schizocarps that can be separated into two mericarp seeds (Palani-Vasantha et al. 2024), and the use of separated mericarps in this study may have allowed more rapid water uptake than would occur in intact schizocarps. However, sufficient water uptake within 4 h does not necessarily imply adequate leaching of germination-inhibiting compound. Therefore, additional soaking time beyond the initial hydration phase is likely required to improve the germination of natural coriander seeds.
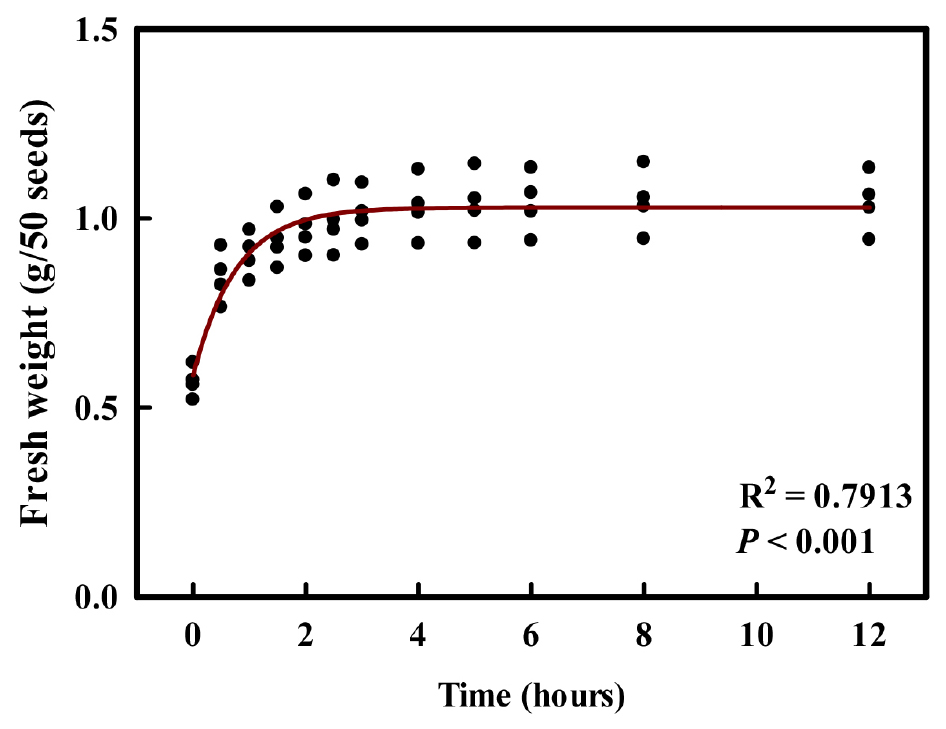
2. Germination parameters
Uneven seed germination and seedling establishment can negatively affect plant uniformity and productivity at later developmental stages (Liu et al. 2019). Therefore, achieving uniform seedling growth at the early stage is important for improving the overall efficiency and productivity of the cultivation process (Kozai et al. 2019).
In this study, the combined effects of temperature and light on coriander seed germination were evaluated. After 10 days of observation, the final germination rate was highest at 15 and 20°C (86.5-90.5%), regardless of light conditions (Fig. 2). Final germination was lowest at 30°C, ranging from only 1.0 to 7.0%. At 10°C, germination progressed slowly during the early period but increased rapidly between days 7 to 9, reaching in a final germination rate of 79.0 to 87.0% (Fig. 2A and B). Nevertheless, cotyledon and radicle development were poor at this temperature. Under dark conditions, germination at 15, 20, and 25°C exceeded that under light conditions by 1.6-, 1.3-, and 1.6-fold, respectively, on day 4, and by 1.1-, 1.1-, and 1.7-fold, respectively, on day 5. In particular, at 25°C, the final germination rate differed by nearly 30% between light and dark treatments, reaching 43.0 and 73.5%, respectively. Regardless of temperature, seedlings germinated under dark conditions exhibited chlorosis in the cotyledons compared with those under light condition (Fig. 2C and D).
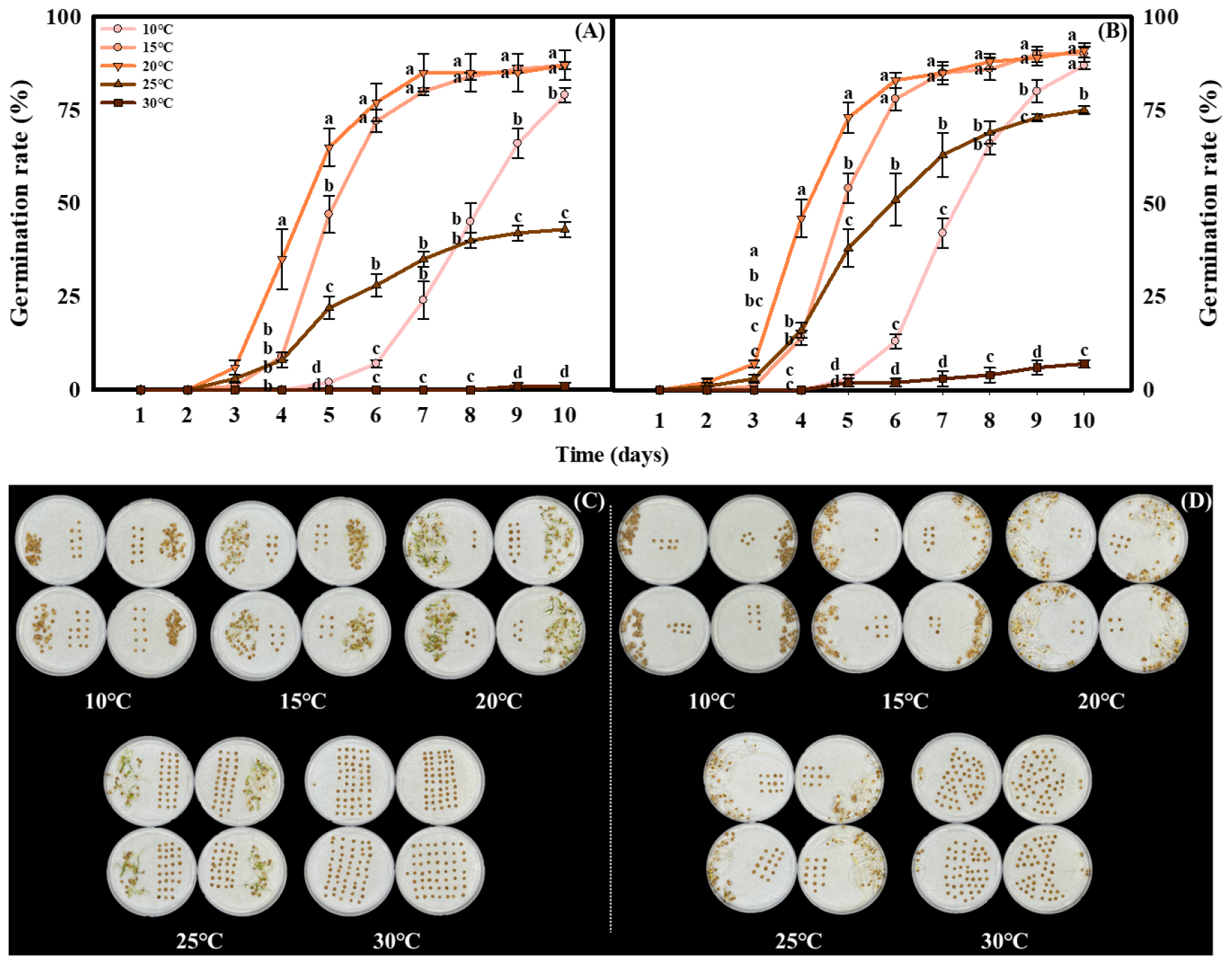
Fig. 2
Germination rate of coriander seeds under light (A) and dark (B) conditions at five different temperatures (10, 15, 20, 25, and 30°C) for 10 days. Error bars indicate standard errors of mean values (n=4). Different letters indicate significant difference at P < 0.001 by Duncan’s multiple range test. Germination of coriander seeds at different temperatures at 10 days after sowing under light (C) and dark (D) conditions
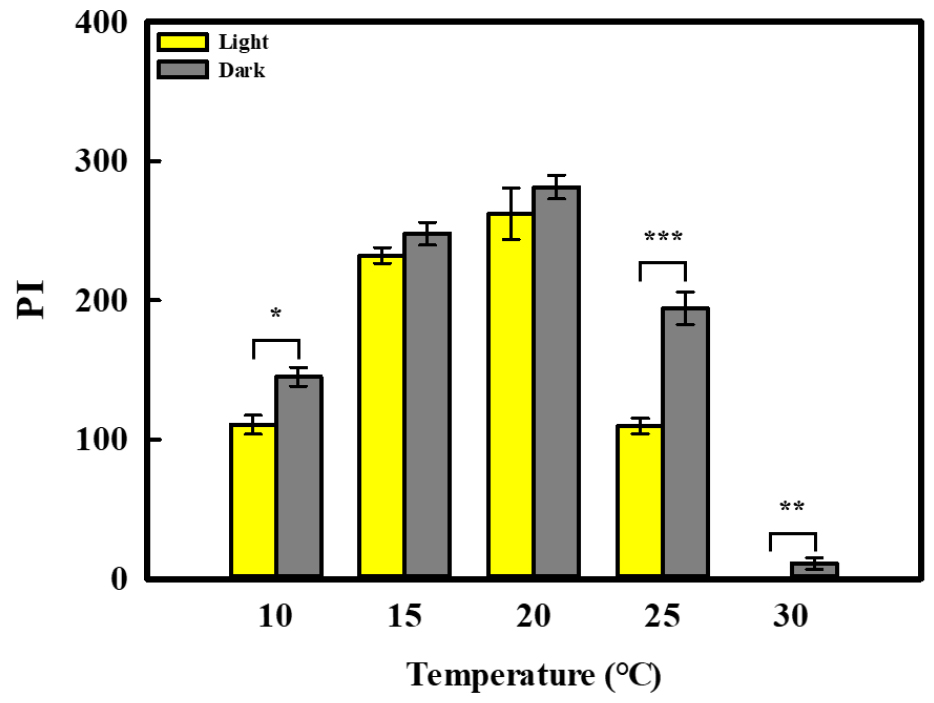
MGT, T50, and PI were calculated from the germination rate data. Regardless of light conditions, MGT was longest at 10 and 30℃, averaging 7.9 and 8.6 days, respectively. At the other temperatures, MGT was shortest at 20℃ and shorter at 15℃ than at 25℃, with the minimum value of 4.9 days observed at 20℃ (Fig. 3A and B). Compared with light conditions, MGT under dark conditions was reduced by an average of approximately 0.4 days, and by approximately 0.2 days at 20°C. T50 could not be determined at 25℃ under light and at 30℃ under either light or dark conditions because the germination rate did not reach 50%. Among the calculated values, T50 was longest at 10℃ (7.4 days) and shortest at 20℃ (4.2 days) (Fig. 3C and D). In contrast, T50 could be estimated at 25℃ under dark because germination was relatively higher than under light condition. Overall, T50 tended to be approximately 0.3 days shorter under dark than under light conditions. PI was lowest at 30°C regardless of light conditions. For the remaining temperature treatments, PI followed the order of 20°C > 15°C > 25°C > 10°C, and the highest value was observed at 20°C (Fig. 4). Light condition did not significantly affect PI at 15 or 20°C, whereas PI was significantly greater under dark conditions at 10, 25, and 30°C.
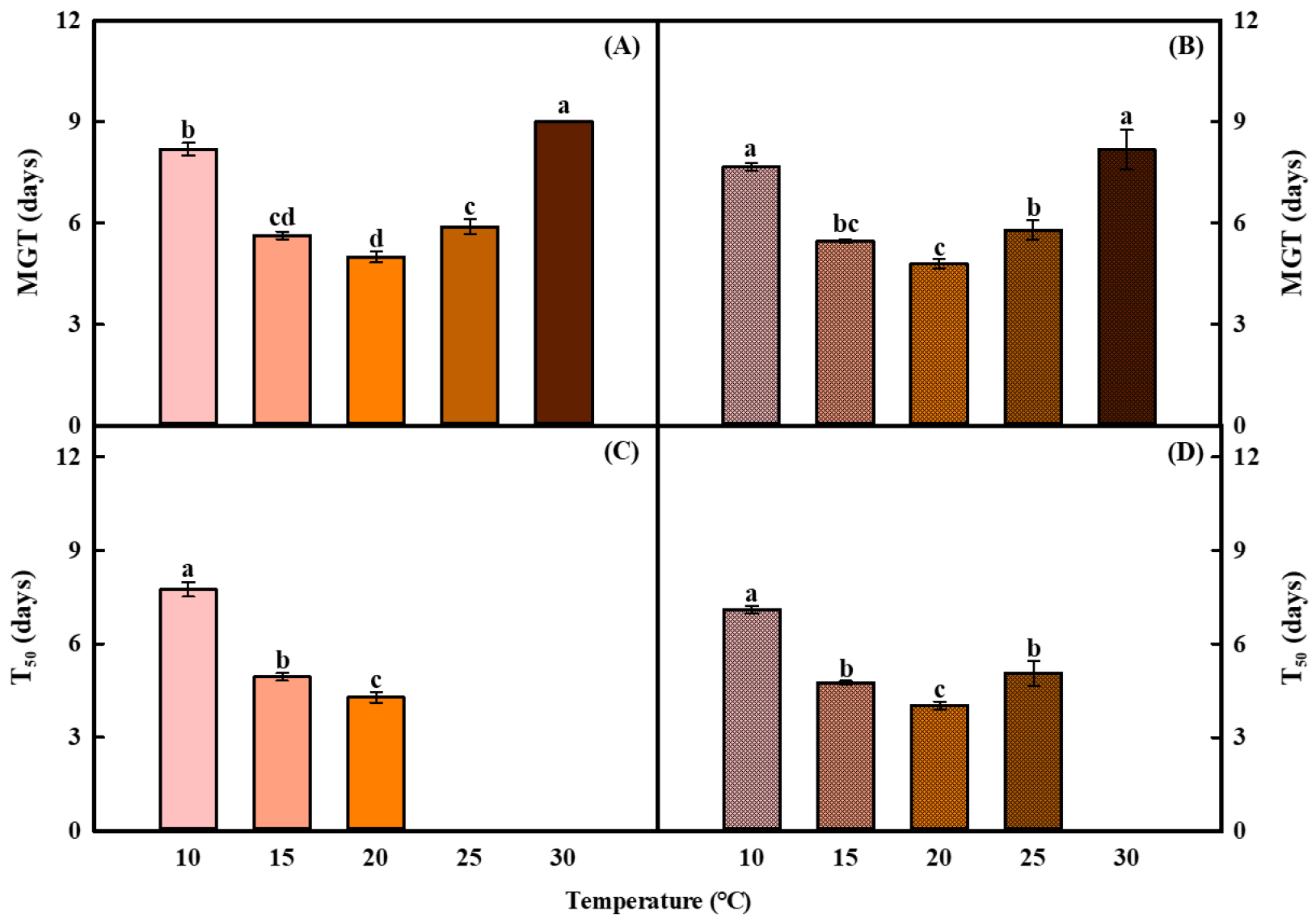
Fig. 3
Mean germination time (MGT) and time to 50% of germination (T50) of coriander seeds under light (A and C) and dark (B and D) conditions at different temperatures (10, 15, 20, 25, and 30°C). T50 at 25 and 30°C under light and at 30°C under dark condition could not be calculated because germination did not reach 50%. Error bars indicate the standard errors of the mean values (n=4). Different letters indicate significant difference at P < 0.001 by Duncan’s multiple range test
Lee et al. (2015) similarly reported that the highest germination rates of coriander seeds occurred at 15 and 20℃ regardless of storage method, whereas temperatures above 25℃ substantial reduced germination. Allahmoradi et al. (2013) also reported high germination rates at 20 and 25°C and estimated the optimal germination temperature to be approximately 24.83°C based on linear regression analysis. Taken together, these findings indicate that coriander seed germination is promoted within temperature ranges similar to those reported previously, although the exact optimum temperature differs somewhat among studies. This suggests that germination responses may vary depending on cultivar, seed condition, and environmental factors other than temperature.
Batlla and Benech-Arnold (2015) reported that temperature primarily regulates germination speed within the permissive temperature range. In the present study, although coriander seeds at 10℃ showed high final germination rates (79% under light condition and 87% under dark condition), germination was initiated much later, resulting in a longer MGT and higher T50. This suggests that, while 10℃ remained within the permissible range for germination and allowed sufficient water uptake and enzymatic activation for radicle emergence, it likely suppressed metabolic processes associated with germination, including cell division and elongation (Zhu et al. 2015). Furthermore, germination at 15 and 20℃ occurred within the optimal temperature range and produced the highest germination rates. In particular, the 20℃ treatment showed the most favorable values for MGT, T50, and PI. This may be attributed to enhanced enzymatic activity and metabolic processes at 20℃, which likely promote ATP production and protein synthesis, thereby accelerating germination (Finch-Savage and Leubner-Metzger 2006; Haj et al. 2022; Khaeim et al. 2022). In contrast, germination rate at 30℃ was nearly absent, which is consistent with the findings of Lee et al. (2015) and Allahmoradi et al. (2013). Generally, temperatures above 30℃ are known to inhibit germination by suppressing enzymatic activity and increasing the respiration rate during germination (Hills and Van 2003; Chiu 2017). Therefore, temperatures above 30℃ appear to be non-permissive for coriander seed germination.
Coriander seeds germinated under both light and dark conditions, indicating that they are not strictly light- dependent and that germination is more strongly influenced by temperature than by light. Nevertheless, dark conditions resulted in superior performance across all germination parameters, with the most pronounced difference observed in PI, an index that places greater weight on early germination. The higher PI under dark conditions indicates that germination proceeded more rapidly during the initial phase. This response may be explained by the more stable microenvironment provided in darkness, including reduced minor temperature fluctuations and more stable oxygen availability for seed respiration (Washa 2015). Additionally, darkness may also serve as a signal of a favorable or safe environment for coriander seed germination (Fenner and Thompson 2005).
Conclusion
Coriander seed germination was strongly influenced by the combination of temperature and light conditions. At 10℃, cotyledon and radicle development were poor, whereas at 30℃, germination was markedly inhibited by high-temperature stress, indicating that both temperatures are unsuitable for coriander seed germination. While germination was most efficient at 20℃ under dark condition, continuous darkness suppresses de-etiolation and negatively affects normal seedling development. Therefore, light exposure at an appropriate stage after radicle emergence is required for proper seedling establishment. In conclusion, these results indicate that maintaining seeds in darkness at 20°C for 4 to 5 days after sowing, followed by light exposure upon radicle emergence, is an effective strategy for producing uniform and vigorous coriander seedlings. This approach is expected to ensure high germination rate and uniform seedling establishment in vertical farming systems.
