

Introduction
Photoperiod and temperature are two main factors affecting the floral induction and runner production in strawberry plants (Durner et al., 1984;Piringer and Scott, 1964). Besides, the application of extra nitrogen (Kim et al., 2013), restricted water supply, or exogenous plant growth regulators (PGRs) can also influence the timing of flowering (Guttridge, 1985;Guttridge and Thompson, 1963;Kim et al., 2017).
The PGRs that are capable of inducing flowering in plants are limited and different from species to species. Among them, cytokinins (Gupta and Maheshwari, 1970;Maheshwari and Venkataraman, 1966), gibberellins (Heide et al., 2013), and salicylic acid (Raskin, 1992) are all documented to induce flowering in some particular species. Benzyladenine (BA), gibberellic acid (GA3), and salicylic acid (SA) are endogenous PGRs of a phenolic nature. They stimulate cell division and expansion, and control and modulate the initiation and development of shoots and roots.
The BA is a synthetic cytokinin that stimulates cell division in plants, promotes vegetative growth, stimulates flowering in some species, and improves fruit quality (Newton and Runkle, 2015). In previous studies, exogenous application of BA proved to be useful for increasing the number of inflorescences and flowers per inflorescence of orchids (Blanchard and Runkle, 2008;Kim et al., 2000;Lee et al., 1998).
The GA3 has various effects on plant growth and development. It can stimulate stem and root growth, induce mitotic division in the leaves of some plants, and increase seed germination rate (Guttridge and Thompson, 1963). In short day plants, GA3 takes part in many physiological pro-cesses such as increasing vegetative development, inhibiting flower bud formation, and enhancing runner growth (Porlingis and Boynton, 1961;Thompson and Guttridge, 1959). Moreover, GA3 at a certain concentration could cause the internode elongation of the main stem, which is rarely induced by the effect of day length.
The SA is an endogenous PGR which plays a role as a natural inductor of thermogenesis, induces flowering in many plant species, and regulates ion uptake and stomatal conductivity (Raskin, 1992). The flowering induction capability of SA has been well known for decades. The effect of SA on this aspect was first reported in a tobacco tissue culture added with indole acetic acid and kinetin (Lee and Skoog, 1965). Other studies have demonstrated that flowering in many plant species has involved SA, such as stimulated flowering in several genera of the Lemnaceae family (Khurana and Cleland, 1992), restored flowering capability in the nutrient-stressed short day species Pharbitis nil (Wada and Takeno, 2010;Wada et al., 2010), and increased numbers of flowers in Gloxinia (Martín et al., 2003) and Calendula officinalis (Pacheco et al., 2013). Although SA can influence flowering in the short day plants, and some long day plant species, its effect still depends on photoperiod, and therefore, selection of the proper daylength might be crucial for induction of flowering (Cleland and Tanaka, 1979).
Strawberry (Fragaria × ananassa) is a perennial stoloniferous plant belonging to the family Rosaceae. With the highly desirable taste and a rich source of phytochemicals (Perez et al., 1997), strawberry is one of the most widely cultivated and consumed fruits in the world (Zobayer et al., 2011). Numerous studies have been published regarding cultivation conditions. The environmental factors and their interactions affect the growth, development, and flowering of strawberry plants. In this study, the effect of exogenous applications of PGRs such as BA, GA3, and SA on growth and flower bud induction at the vegetative growth stage in strawberry ‘Sulhyang’ was investigated.
Materials and Methods
1. Plant materials and exogenous applications of plant growth regulators
Runner plants of strawberry ‘Sulhyang’ were collected from a strawberry farm (Sugok-myeon, Jinju, Gyeongsangnam- do, Korea) and rooted in 10cm plastic pots filled with a BVB medium (Bas Van Buuren Substrates, EN- 12580, De Lier, The Netherlands). These runner plants were irrigated daily with a greenhouse multipurpose nutrient solution [in mg·L-1 Ca(NO3)2·4H2O 737.0, KNO3 343.4, KH2PO4 163.2, K2SO4 43.5, MgSO4·H2O 246.0, NH4NO3 80.0, Fe-EDTA 15.0, H3BO3 1.40, NaMoO4·2H2O 0.12, MnSO4·4H2O 2.10, and ZnSO4·7H2O 0.44 and an electrical conductivity 0.8 mS·cm-1].
After 3 weeks, the plants were placed in growth chambers and the culture condition was set at 25°C/12°C (day/ night) temperature, the light intensity of 300μmol·m-2·s-1 PPFD, and 70 ± 5% relative humidity. A 14 h photoperiod (LD, long day) was given at two periods during the experiment, first 3 weeks and from week 7 to the end of the experiment. During weeks 4 to 6, a 10 h photoperiod (SD, short day) was given (Fig. 1).
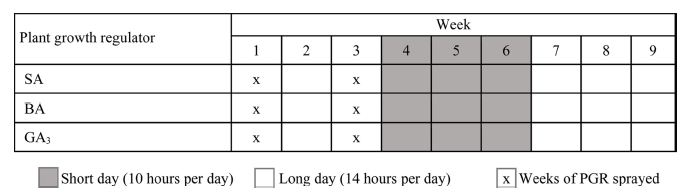
The runner plants were treated with one of 3 concentrations, 0 (control), 100, and 200mg·L-1 of BA, GA3, or SA solution. These solutions were applied by foliar spray for two times, each time with 10mL per plant, at an interval of 2 weeks (Fig. 1).
2. Measurements of growth and morphological parameters
After 9 weeks, shoot length (from the crown to longest leaf tip), root length, leaf area, chlorophyll index (SPAD), number of runners, runner fresh weight, and plant fresh and dry weights were measured. Leaf area was taken from all leaves detached by using a leaf area meter (LI-3100, LICOR Inc., Lincoln, NE, USA). Chlorophyll was measured using a chlorophyll meter (SPAD-502 Plus, Konica Minolta Sensing Inc., Osaka, Japan). Dry weight was measured after drying the samples for 72 hours in a dry oven (FO- 450M, Jeio Technology Co., Ltd., Seoul, Korea) at 80°C.
3. Statistical analysis
The experiment was set up in a completely randomized design with 6 plants per treatment. Data collected were analyzed for statistical significance by the SAS (Statistical Analysis System, V. 9.1, Cary, NC, USA) program. The experimental results were subjected to an analysis of variance (ANOVA) and Duncan’s multiple range tests. Graphing was performed with OriginPro 9.0 (OriginLab Corporation, Northampton, MA, USA).
Results and Discussion
The BA, GA3, and SA are plant growth regulators that play important roles in controlling growth and development of plants (Beyl, 2016;Rivas-San Vicente and Plasencia, 2011). After cultivation for 9 weeks in the control environment, the effect of type and their concentrations of PGRs on the vegetative growth of strawberry plants was significantly different (Fig. 2). The enhancement of shoot length was observed on the plants that were treated with all PGRs at a concentration of 100mg·L-1 (Table 1). The greatest shoot length (27.6cm) was recorded in the GA3 treatment, followed by BA and SA at a concentration of 100mg·L-1. The higher concentrations of these PGRs caused a reduction in shoot length. The shoot length of plants treated with a concentration of 200mg·L-1 was 3-6cm shorter as compared to that of plants treated with a lower concentration (Table 1). There were no significant differences in root length among the treatments. However, the application of PGRs in this study caused a reduction in root length as compared to the control, and the greatest root length (14.5cm) was obtained in the control. In all treated plants, the leaf area was considerably larger than that in the control, especially in a higher concentration (200mg·L-1) of PGRs (Table 1). At the same level of PGRs, the greatest leaf area was obtained in the GA3 treatment, followed by SA and BA. Nonetheless, the SPAD index of chlorophyll showed that the application of BA or GA3 to the plants at both concentrations caused the decrease in chlorophyll content (Table 1). The leaf area of the plants treated with GA3was enhanced by 56% as compared to the control, and this value in the SA treatment was 25%. Martín et al. (2003) reported that foliar spray in Gloxinia at the concentration of 10-8 M SA resulted in the increase of leaf area by 49%.

Table 1. Effect of benzyladenine (BA), gibberellic acid (GA3), and salicylic acid (SA) on growth and development of strawberry ‘Sulhyang’ after 9 weeks.
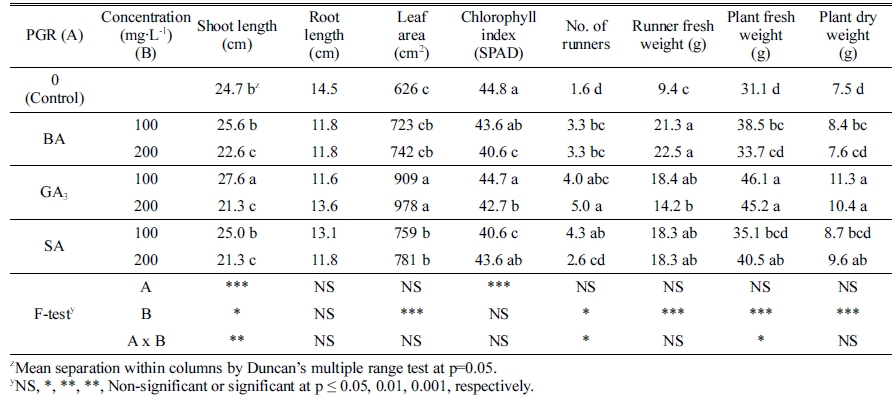 |
The application of those PGRs and their concentrations in this study also affected the formation and development of runners. The number of runners was greatly increased in the PGRs treatments. The greatest number of runners (5.0 runners per plant) was observed in the treatment of 200mg·L-1 GA3 as compared to that in plants treated with BA (3.3 runners per plant) and SA (2.6 runners per plant). Interestingly, the number of runners in plants treated with BA or GA3 increased with the increase in their concentration (Table 1). A similar finding by Thompson and Guttridge (1959) reported that the number of runners per strawberry crown increased with the increase in GA3 concentration. On the other hand, the increase in the concentration of SA resulted in decrease in the number of runners. Weight of the runners separated from the mother plants after 9 weeks of cultivation, showed that the BA strongly enhanced the runner fresh weight, either at low or high concentration, by at least two times as compared to the control. However, an abnormal phenomenon was observed in the plants treated with GA3. The first effects of those PGRs on strawberry plants appeared at about two weeks after the treatment with PGRs. The youngest leaf of the treated plants was slightly paler and larger as compared to that of other treatments. The internodes of mother plants and new daughter plants in these plants were oddly elongated (Figs. 2 and 3). Kang et al. (2018) and Thompson and Guttridge (1959) also observed the same phenomenon in strawberry plant treated with GA3. This phenomenon may cause some troubles in utilizing and growing of the daughter plants as well as the mother plants.

Fresh weight of strawberry plants was significantly affected by PGRs treatments combined with their concentrations (Table 1). The greatest plant fresh weight was observed in the treatment of GA3 regardless of its concentration. Plant fresh weight was 46.1 and 45.2g per plant in the treatment with GA3 at a concentration of 100 and 200mg·L-1, respectively, as compared to the plant in the control treatment having only 31.1 g per plant. Plant dry weight showed the same trend as the fresh weight (Table 1). In the agreement with this results, a previous study demonstrated that the application of 100mg·L-1 GA3 on strawberry plants only promoted in the initiation of runners, shoot length and petiole length, but not flower initiation (Thompson and Guttridge, 1959). In the long day condition, the vegetative growth of strawberry plants, such as the development of runners in the leaf axils, elongation of petiole and upright growth, was remarkably enhanced. Therefore, GA3 would be considered as a substitute for a growth-promoting hormone which was typically produced naturally under long day condition in strawberry plants.
The flowering plants and number of inflorescences per plant were only observed in the treatment of SA. They were increased with the increase of SA concentration (Fig. 4). Results showed that up to 85% of plants treated with 200mg·L-1 SA flowered after 6 weeks from the beginning of short day period. However, only 58% of plants flowered in the treatment of 100mg·L-1 SA and there were no flowers observed in the other treatments. The number of inflorescences also showed the same trend. The greatest number of inflorescences (4.3 inflorescences per plant) was obtained in the 200mg·L-1 SA, followed by 100mg·L-1 SA (2.3 inflorescences per plant). Similar results were also observed in Lemna gibba G3, in which the exogenous application of SA dramatically induced flowering while having a slight effect on subsequent development of flowers (Cleland and Tanaka, 1979;Conn, 1984). The mechanism of flower induction effect of SA in the plant is mysterious. Research suggested that to induce flowers, SA might behave as a chelating agent in the plant (Oota, 1975). And this hypothesis has been reinforced by the fact that chelating agents could induce flowering in Lemnaceae (Oota, 1972;Seth et al., 1970).
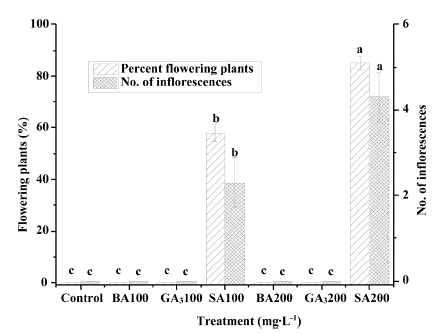
Fig. 4
Effect of concentration of foliar-sprayed benzyladenine (BA), gibberellic acid (GA3), and salicylic acid (SA) on the number of flowers in strawberry ‘Sulhyang’ measured after 30 days. Means accompanied by different letters are significantly different (p < 0.05) according to the Duncan’s multiple range test. Vertical bars indicate the standard error (n=6).
Overall, the results indicated that the application of BA, GA3, or SA could change various growth traits in strawberry plants. In particular plant species, BA displayed as the ideal factor for increasing percentage plants with inflorescence, and inducing earlier flowering. However, the present study demonstrated that the BA did not have the same effect in strawberry plants tested. Vegetative growth of strawberry plants treated with GA3 was significantly enhanced. However, the abnormal elongation of internodes of main stem and daughter plants with the 200mg·L-1 GA3 could be disadvantages. The recommended concentration of GA3 would be 100mg·L-1 accounting both normal development and high growth traits. It should be noted that the GA3 accelerated the vegetative growth, and consequently the flowering time may be delayed. As expected, SA played an important role in increasing the number of inflorescences in strawberry plants. At a concentration of 200mg·L-1 SA, not only the number of inflorescences, but also many growth traits were virtually enhanced as compared to the control. However, because of the limitation in time of this study, the yield and fruit quality were not assessed. A possibility may arise that despite hastening in flowering in the SA treatment, more vigorous plants in the GA3 treatment may induce more number of flowers to give increased fruit yield. Therefore, further studies are still needed, especially, to reveal the hypothesis that the combination of GA3 and SA may improve by not only increased vegetative growth, but also shortened time to flower induction.
