

Introduction
Materials and Methods
1. Planting material and experimental site
2. Cultivation conditions
3. Growth indices measurement
4. Nutrient solution composition
5. Inorganic nutrient analysis
6. Statistical analysis
Results and Discussion
1. Growth response of leaf perilla to phosphorus deficiency
2. Effects of phosphorus deficiency (PD) on mineral nutrient accumulation
3. Symptoms of leaf perilla induced by phosphorus deficiency
Introduction
Perilla (Perilla frutescens Britt.), an annual herbaceous plant belonging to the Lamiaceae family, is traditionally cultivated in East Asian countries, including Korea, China, Japan, and India (RDA, 2018). In Korea, the cultivation of perilla leaves holds significant economic value, with an estimated cultivation area of approximately 1,100 ha, an annual production of about 40,000 tons, and a market value of around 220 billion KRW as of 2020 (MAFRA, 2022).
Perilla leaves are highly valued for their nutritional and functional properties, being rich in calcium, essential minerals, vitamins, and rosmarinic acid, a potent antioxidant. These attributes have led to increased recognition of perilla as a health-promoting food ingredient. In response to growing consumer demand, exports to neighboring countries, particularly Japan, have steadily increased (Kim et al., 2021).
Traditionally, perilla has been cultivated in open-field or soil-based greenhouse systems. However, repeated cultivation in the same soil has led to the accumulation of soilborne pathogens and the emergence of physiological disorders, posing challenges to sustainable production. To overcome such limitations, hydroponic cultivation systems have emerged as a promising alternative, offering precise control over nutrient delivery and root-zone conditions. These systems not only improve crop yield and quality but also enhance resource use efficiency and reduce labor requirements (Dorais et al., 2001; Nam et al., 2019). Moreover, hydroponics help mitigate issues associated with soilborne diseases and nutrient imbalances, contributing to environmentally sustainable agricultural practices.
Among various hydroponic methods, substrate-based systems utilizing materials such as coir and rockwool currently dominate commercial operations, accounting for more than 90% of hydroponic cultivation in Korea (MAFRA, 2022). However, long-term use of these substrates can lead to physical and chemical degradation, including reduced porosity, waterlogging, and difficulty in maintaining optimal root-zone temperatures (RDA, 2021). These limitations have spurred growing interest in alternative systems such as the Deep Flow Technique (DFT), which provides a continuous supply of water and nutrients through a recirculating solution and is particularly suitable for green leafy vegetable production. Despite its potential, research on the application of DFT to perilla remains limited, as most existing studies have focused on substrate-based hydroponics (Kim et al., 2023; Shin et al., 2022).
Optimal nutrient management is essential in hydroponic systems, where plant growth, physiological responses, and product quality are directly influenced by nutrient availability. Among essential mineral elements, phosphorus (P) plays a pivotal role in energy transfer, nucleic acid synthesis, and membrane formation (Marschner, 2012). Phosphorus deficiency can lead to significant growth inhibition, reduced biomass accumulation, and visible deficiency symptoms (Bennett, 1993; Bould et al., 1983; Nelson, 2003). In hydroponics, where nutrient supply is entirely dependent on solution composition, it is crucial to understand crop-specific requirements and physiological responses to nutrient imbalances in order to develop precise and efficient fertilization strategies (Yamazaki, 1982).
Therefore, the objective of this study was to evaluate the effects of phosphorus deficiency on the growth and physiological characteristics of perilla cultivated under Deep Flow Technique (DFT) conditions. By systematically adjusting phosphate concentrations in the hydroponic solution, this research aims to elucidate the growth responses of perilla, thereby contributing to the development of optimized nutrient management strategies for sustainable and high-efficiency hydroponic perilla production.
Materials and Methods
1. Planting material and experimental site
The experiment was conducted using the leaf perilla cultivar ‘Namcheon’ (Perilla frutescens Britt. cv. ‘Namcheon’). Seeds were sown on February 24, 2023, in 200-cell plug trays filled with commercial nursery substrate. After germination, seedlings of two weeks age that had developed at least two fully expanded true leaves were selected for uniformity. Prior to transplantation, seedling roots were thoroughly rinsed to remove residual substrate. Each plant was then inserted into a cubical hydroponic sponge with a central hole, specifically designed to support seedlings, and placed into a simplified hydroponic cultivation system (Higreen Hydroponic Machine B, Gawhatech, Korea). The experiment was conducted in a 40 m2 Venlo-type controlled glass greenhouse located at Pusan National University, which maintained consistent environmental conditions throughout the cultivation period.
2. Cultivation conditions
Uniform seedlings were selected and transplanted into a deep flow technique (DFT) hydroponic system. Each hydroponic unit was filled with 10 L of nutrient solution. The nutrient solution was completely renewed every two weeks during the first 40 days, and weekly thereafter. The greenhouse temperature was maintained at a constant 20°C throughout the day and night. To suppress floral bud initiation during the entire experimental period, 200 W metal halide lamps were installed at 2-meter intervals and suspended 2 meters above the plant canopy to provide photoperiodic lighting from sunset to sunrise. Lateral branches were pruned once or twice per week to maintain the dominance of the main stem. The experimental design consisted of four plants per treatment, with three replicates per treatment.
3. Growth indices measurement
Growth indices, including plant height, number of leaves, number of nodes, and stem diameter, were measured at 20, 30, 50, 60, and 80 days after transplanting. Chlorophyll content was assessed beginning at 30 days after transplanting. At both 60 and 80 days, three plants per treatment were randomly selected to determine fresh and dry weights. Additionally, accumulated leaf area per plant was calculated at 80 days. The number of leaves was recorded by counting leaves longer than 3 cm, and leaf area was measured using a leaf area meter (LI-3000, LI-COR, USA). Stem diameter was measured 1 cm above the shoot base using a digital vernier caliper (DC150P, Bluetec, China). Fresh weight was recorded immediately after harvest, while dry weight was obtained after drying plant samples at 70°C for 72 hours. Chlorophyll content was measured using a SPAD chlorophyll meter (SPAD-502, Minolta Co., Ltd., Japan), with three readings taken per leaf. The number of nodes was determined by counting the leaf-bearing nodes with leaves exceeding 3 cm in length from the shoot base.
4. Nutrient solution composition
The base nutrient solution used in this study was the Yamazaki hydroponic formulation, commonly applied for green leafy vegetables, and served as the control treatment. The composition of the control solution was as follows (in me·L-1): NO--N, 6.0; NH4+-N, 0.5; PO43--P, 1.5; K, 6.0; Ca, 2.0; Mg, 1.0; and SO42--S, 4.0. Micronutrient concentrations (in mg·L-1) were: Fe, 3.0; BO3-, 0.5; Mn, 0.5; Zn, 0.05; Cu, 0.05; and MoO42-, 0.001. Phosphorus-deficient treatments were prepared by modifying the phosphate concentration in the Yamazaki nutrient solution for leafy vegetables. The control treatment (0% deficiency) contained 1.5 me·L-1 of phosphate. Based on this, nutrient solutions were formulated with reduced phosphate concentrations to create 30% deficiency (0.45 me·L-1), 50% deficiency (0.75 me·L-1), 70% deficiency (1.05 me·L-1), and 100% deficiency (0.0 me·L-1).
The electrical conductivity (EC) of each nutrient solution was adjusted to 1.0 dS·m-1 (Nguyen et al., 2021), and the pH was maintained within the range of 6.0 throughout the experiment.
5. Inorganic nutrient analysis
To evaluate the effects of phosphorus deficiency on nutrient uptake, the aerial parts of the plants were analyzed for total nitrogen (T-N), phosphorus (P), potassium (K), calcium (Ca), and magnesium (Mg) content. Dried plant tissues were initially ground using a Wiley mill (Thomas-Wiley Mill Co., Philadelphia, PA, USA) to pass through a 1 mm mesh screen, followed by fine grinding with a cyclone impact mill (Udy Corporation, Fort Collins, CO, USA) to ensure sample homogeneity. For T-N analysis, 0.3 g of ground tissue was digested with concentrated sulfuric acid (H2SO4), and nitrogen content was determined using a semi-automatic Kjeldahl nitrogen analyzer (Pro-Nitro S, Model 4002851, J.P. Selecta, Barcelona, Spain). Phosphorus concentration was measured using the molybdenum blue colorimetric method following acid digestion. Potassium levels were determined via atomic absorption spectrophotometry (AA- 7000, Shimadzu Corporation, Kyoto, Japan), while calcium and magnesium concentrations were analyzed using EDTA complexometric titration. Each nutrient analysis was performed in triplicate to ensure data reliability and precision.
6. Statistical analysis
Statistical analysis was conducted using SAS software (version 9.4; SAS Institute Inc., Cary, NC, USA). Treatment effects were evaluated using Duncan’s Multiple Range Test (DMRT) at a significance level of p ≤ 0.05.
Results and Discussion
1. Growth response of leaf perilla to phosphorus deficiency
Phosphorus (P) is an essential macronutrient involved in various physiological and biochemical processes, including energy transfer, membrane formation, and nucleic acid synthesis (Marschner, 1995; Mengel and Kirkby, 1987). The results of this study demonstrate a significant growth- suppressing effect of phosphorus deficiency on leaf perilla under hydroponic conditions.
Low phosphorus deficient groups (30%, 50%, 70% ‘P’ deficiency) have shown a moderate reduction in all growth parameters, while 100% phosphorus deficient group has an abrupt decline in growth along with statistically significant difference to that of control. This trend persisted throughout the growth period at 20, 30, 50, 60, 80 days after transplanting. At 80 days growth stage, the decline in various growth parameters was as follows: Plant height by 86% (73.5 cm vs 10.1 cm), number of leaves by 78% (12.8 leaves vs 2.7 leaves), number of nodes by 72% (10.8 nodes vs 3.0 nodes), stem diameter by 69% (10.9 mm vs 3.3 mm) compared to the control group (Table 1, Fig. 3). This supports the previous findings stating that phosphorus deficiency limits the vegetative development (Choi and Park, 2007).
Table 1.
Effects of phosphorus deficiency on plant height, leaf number, node number, and stem diameter in hydroponically grown of ‘Namcheon’ perilla.
| P deficiencyz (%) |
Plant height (cm) | No. of leaves | No. of nodes |
Stem diameter (mm) |
| 20 days after treatment | ||||
| 0y | 7.9 abx | 5.2 a | 2.6 a | 3.5 a |
| 30 | 9.4 a | 5.0 a | 2.5 a | 3.5 a |
| 50 | 8.6 a | 5.0 a | 2.5 a | 3.3 a |
| 70 | 9.6 a | 5.7 a | 2.8 a | 3.4 a |
| 100 | 6.1 b | 4.0 b | 2.0 b | 2.6 b |
| 30 days after treatment | ||||
| 0 | 23.4 a | 7.7 a | 4.8 ab | 7.0 a |
| 30 | 25.0 a | 8.0 a | 5.0 a | 6.9 a |
| 50 | 21.9 a | 7.7 a | 4.8 ab | 6.9 a |
| 70 | 23.9 a | 7.2 a | 4.6 b | 7.2 a |
| 100 | 9.8 b | 4.7 b | 2.1 c | 2.6 b |
| 50 days after treatment | ||||
| 0 | 36.6 a | 6.5 a | 7.3 a | 8.5 ab |
| 30 | 36.1 a | 7.0 a | 6.5 b | 8.7 ab |
| 50 | 33.8 a | 6.7 a | 6.3 b | 8.9 a |
| 70 | 35.3 a | 6.7 a | 6.3 b | 8.2 b |
| 100 | 8.9 b | 4.0 b | 2.3 c | 2.8 c |
| 60 days after treatment | ||||
| 0 | 56.2 a | 10.8 a | 8.2 a | 9.6 a |
| 30 | 48.8 ab | 9.0 a | 7.6 a | 9.1 a |
| 50 | 47.2 ab | 9.3 ab | 7.0 ab | 9.2 a |
| 70 | 56.0 a | 9.7 a | 7.7 a | 9.2 a |
| 100 | 9.3 b | 4.0 b | 2.5 b | 3.0 b |
| 80 days after treatment | ||||
| 0 | 73.5 a | 12.8 a | 10.8 a | 10.9 a |
| 30 | 69.2 a | 10.0 ab | 10.3 a | 10.1 a |
| 50 | 65.2 a | 10.0 ab | 9.5 ab | 10.1 a |
| 70 | 65.1 a | 10.0 ab | 8.3 bc | 9.8 a |
| 100 | 10.1 b | 2.7 c | 3.0 c | 3.3 b |
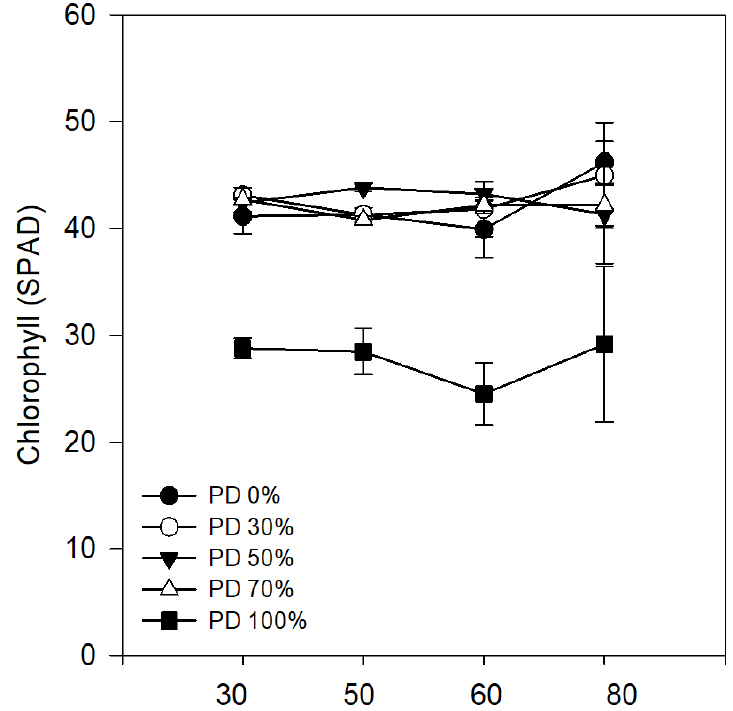
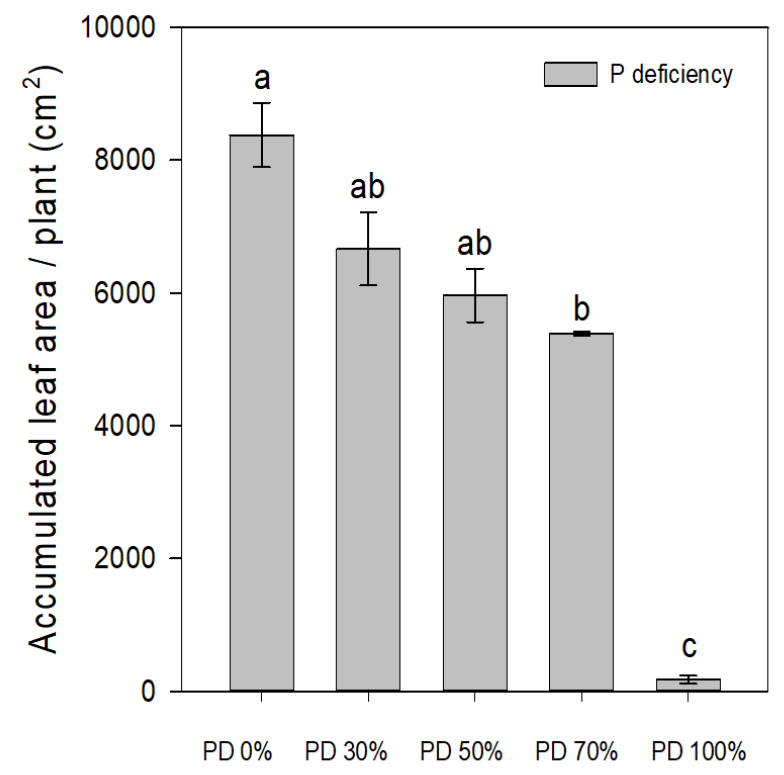
Similarly, at 80 days total plant fresh weight and dry weight has exhibited a more severe suppression in 100% phosphorus deficient group. Total plant fresh weight and dry weight has been suppressed by 97% (93.4 g vs 2.1 g) and 96% (11.8 g vs 0.4 g) respectively indicating the drastic inhibition of biomass accumulation (Table 2). These results confirm earlier reports concerning the positive co-relation of phosphorus availability with biomass production (Choi et al., 2009). SPAD values, indicative of chlorophyll content, were significantly lower under complete phosphorus deficiency, ranging from 24.5 to 29.2, in contrast to 40.8-46.2 observed in the other treatments (Fig. 1). These results suggest impaired chlorophyll biosynthesis under phosphorus- limited conditions, consistent with earlier findings (Choi and Park, 2007). Leaf area exhibited a clear phosphorus concentration-dependent decline. By day 80, plants in the control group achieved a total leaf area of 8,986.4 cm2, while those subjected to complete phosphorus deprivation (100% deficiency) showed a pronounced reduction to just 210.6 cm2 (Fig. 2). These findings highlight the indispensable role of phosphorus in maintaining optimal leaf expansion, and by extension, vegetative growth, chlorophyll content, and overall photosynthetic capacity in Perilla frutescens.
Table 2.
Effects of phosphorus deficiency on fresh weight and dry weight in hydroponically grown of ‘Namcheon’ perilla.
| P deficiencyz (%) | Fresh weight (g/plant) | Dry weight (g/plant) | ||||
| Leaves | Stem | Total | Leaves | Stem | Total | |
| 60 days after treatment | ||||||
| 0y | 60.3 ax | 25.8 a | 86.1 a | 5.0 a | 3.7 a | 8.7 a |
| 30 | 52.5 ab | 25.0 ab | 77.5 a | 4.6 a | 3.4 a | 8.0 a |
| 50 | 47.1 ab | 18.8 ab | 65.8 a | 4.3 a | 3.2 a | 7.5 a |
| 70 | 44.8 b | 22.0 ab | 66.8 a | 3.8 a | 3.8 a | 7.6 a |
| 100 | 1.2 c | 1.1 b | 2.3 b | 0.6 b | 0.3 b | 0.9 b |
| 80 days after treatment | ||||||
| 0 | 62.4 a | 31.1 a | 93.4 a | 7.0 a | 4.8 a | 11.8 a |
| 30 | 57.2 ab | 34.5 a | 91.7 ab | 5.5 a | 4.7 a | 10.2 a |
| 50 | 50.2 b | 26.5 ab | 76.7 c | 6.4 a | 4.3 a | 10.7 a |
| 70 | 52.9 ab | 25.5 ab | 78.4 bc | 5.3 a | 4.8 a | 10.1 a |
| 100 | 0.8 c | 1.3 b | 2.1 d | 0.1 b | 0.3 b | 0.4 b |

2. Effects of phosphorus deficiency (PD) on mineral nutrient accumulation
To investigate the influence of phosphorus availability on nutrient composition, the aerial tissues of Perilla frutescens were observed after 80 days of treatment. The data showed a clear, concentration-dependent decline in tissue phosphorus content with increasing concentration of external phosphorus deficiency (Table 3, Fig. 4).
Table 3.
Mineral element composition of ‘Namcheon’ perilla after 80 days of hydroponic cultivation with Yamazaki nutrient solution based on three replicates.
| T-N | P2O5 | K2O | CaO | MgO |
| (%) | (%) | (%) | (%) | (%) |
| 2.9 ± 0.03 | 1.8 ± 0.03 | 2.7 ± 0.21 | 2.0 ± 0.11 | 0.6 ± 0.01 |
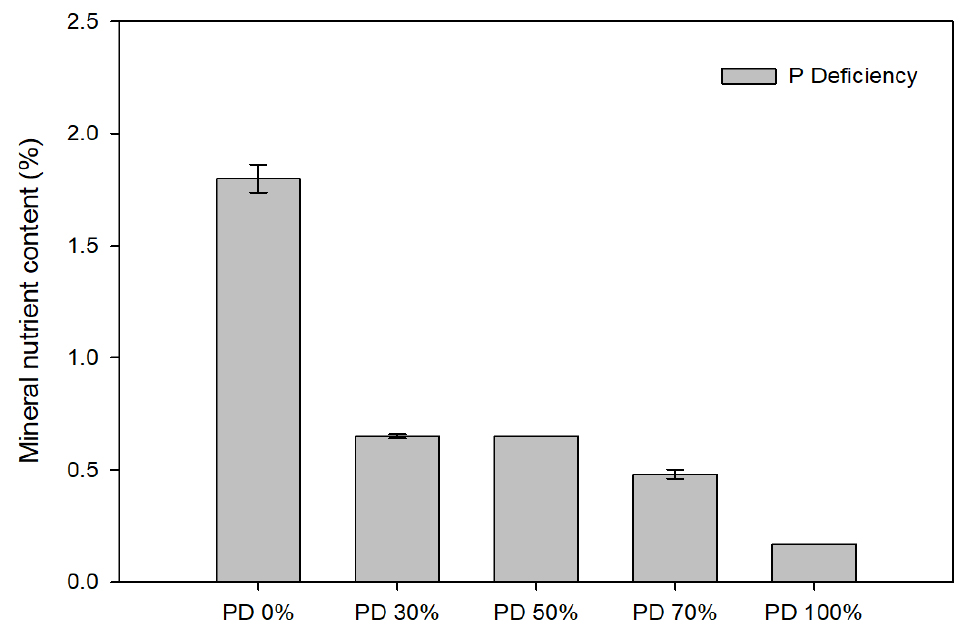
In the control treatment (PD 0%), tissue phosphorus concentration was 1.80%, which decreased markedly to 0.65% under both PD 30% and PD 50% treatments. A further reduction to 0.48% was observed in the PD 70% treatment. The most dramatic decline was recorded in the PD 100% treatment, where phosphorus concentration dropped to 0.17%, representing approximately 9% of the control level. This substantial reduction highlights a severe impairment of phosphorus uptake and accumulation under complete phosphorus deprivation, consistent with previous findings (Kim et al., 2003; Marschner, 2012).
In addition to phosphorus, the concentrations of essential cations—potassium (K2O), calcium (CaO), and magnesium (MgO)—were also affected by phosphorus availability. In the control group, the respective concentrations were 2.7% for K2O, 2.0% for CaO, and 0.6% for MgO. Under phosphorus- deficient conditions, all three nutrients exhibited declining trends, with the lowest values recorded in the PD 100% group. These results suggest that phosphorus deficiency may interfere with the uptake or transport of other mineral elements, likely due to disruptions in root physiology under nutrient stress.
The interactions observed between phosphorus and other minerals may be attributed to phosphorus’s regulatory role in root membrane permeability, ATP-dependent ion transport, and exudation processes, as reported in previous studies (Mengel and Kirkby, 1987; Raghothama, 1999). The decline in cation concentrations under phosphorus stress underscores the importance of phosphorus in maintaining mineral nutrient balance and suggests the potential for secondary deficiencies when phosphorus availability is insufficient.
Overall, the findings demonstrate that phosphorus deficiency not only limits its own accumulation but also negatively affects the uptake of other essential nutrients. This ultimately compromises the nutritional status and physiological resilience of the plant, reinforcing the critical role of phosphorus in plant nutrition.
3. Symptoms of leaf perilla induced by phosphorus deficiency
Severe phosphorus deficiency led to pronounced physiological stress in hydroponically cultivated leaf perilla, most notably in the 100% phosphorus-deficient treatment, where no phosphorus was supplied throughout the growing period. Visual symptoms first appeared on older leaves, reflecting the high phloem mobility of phosphorus and its tendency to be remobilized to younger tissues under deficient conditions (RDA, 2018).
Characteristic symptoms included dark brown pigmentation on the adaxial (upper) leaf surface and purplish discoloration on the abaxial (lower) surface, followed by necrosis and premature leaf abscission (Fig. 5). These symptoms are indicative of anthocyanin accumulation and cellular degradation, which are commonly associated with phosphorus starvation (Schachtman et al., 1998). In addition, stem elongation was markedly suppressed in the 100% phosphorus- deficient treatment, with stems becoming thin, brittle, and structurally compromised.

These morphological abnormalities can be attributed to disruptions in key metabolic functions. Phosphorus is essential for ATP synthesis, nucleic acid replication, and phospholipid biosynthesis (Marschner, 2012; Raghothama, 1999). Its deficiency impairs energy transfer, restricts cell division, and weakens membrane integrity—factors that collectively result in stunted growth and increased susceptibility to environmental stressors (Plaxton and Tran, 2011).
Furthermore, phosphorus plays a critical role in regulating photosynthetic enzymes and supports the Calvin cycle by maintaining adequate levels of ATP and NADPH (Foyer and Spencer, 1986). The observed reductions in chlorophyll content and photosynthetic performance under phosphorus- deficient conditions are consistent with these physiological limitations.
In summary, these findings underscore the fundamental importance of phosphorus in sustaining physiological and morphological integrity in leaf perilla. Early detection of visual deficiency symptoms can serve as an effective diagnostic tool for phosphorus status, enabling timely intervention and improved nutrient management in hydroponic systems to support sustainable and high-yield production.
