

서 론
우리나라의 토마토 시설재배면적은 6,976ha이고, 생산 량은 457천톤이며(2016, MAFRA), 시설원예 현대화를 위한 정부의 지원으로 토마토 수경재배 면적은 매년 증 가하고 있다(602.4ha, 1,350농가, 2017, RDA). 수경재배 는 균형 시비, 비료의 흡수효율 증대, 토양전염성 병원 균의 회피, 위생적 재배환경 관리를 통한 연작장해의 감 소, 생산과정의 생력화, 그리고 고품질 농산물 생산과 생산량 증대 등의 다양한 장점을 갖는다.
그러나 대부분의 수경재배 농가에서는 비순환식 수경 재배 방법으로 토마토를 생산하고 있으며, 이 과정에서 공급되는 총 양액 중 20~30%가 배액으로 토양이나 하 천으로 배출되어 지하수와 하천의 오염을 가중시킨다 (RDA, 1999). 또한 배출되는 배액은 물과 비료의 과다 사용을 유발하여 생산비를 증가시키는 원인으로 보고되 고 있다(Bellert et al., 1998; Benoit, 1992; Rhee et al., 2003; Roh et al., 1997; Smith, 1997; Zhang et al., 2010a). van Os(1994)는 토마토 수경재배 시 배액량의 20% 정도가 배출될 경우 ha당 연간 질소 147kg, 인산 71kg, 칼륨 282kg, 마그네슘 60kg의 비료가 손실된다고 보고하였고, Sonneveld(1993)은 비순환식과 비교해 순환 식 수경재배가 물 소비량을 64% 절감하고 질소와 칼륨 소비량을 44~50% 감소할 수 있다고 하였다.
이상의 문제로 인해 수경재배에서 발생된 배출액을 재 활용하기 위해 관비 용액으로 재활용(Zhang et al., 2010b), 배액의 전기충격살균법(Lee et al., 2016), membrane filter를 이용한 배액 재활용 기술 개발(Bernard, 1991; Ohtani et al., 2000) 등의 다양한 연구가 수행되었다. 이 중 배액을 다시 양액으로 재사용하기 위해서는 공급된 양액과 배액에 포함된 각종 무기원소의 농도 변화에 대하 여 정확한 분석 데이터가 있어야 하고, 이러한 정보를 기 반으로 추가할 무기원소의 종류 및 농도가 결정되어야 한 다. 이럴 경우 배액을 재활용하여도 각종 무기원소 농도를 최초 공급하는 시기의 양액 조성과 유사하게 유지할 수 있고, 무기원소 농도 변화에 기인한 수확량 감소를 방지할 수 있다. 그러나 방울토마토 재배 시 이와 관련된 연구결 과가 충분하지 못해 참고할 문헌이 부족하다.
따라서 방울토마토 재배 시 착과 절위에 따른 생육특 성, 공급 배양액과 배출액의 무기원소 농도 변화 그리고 식물체의 무기성분 함량을 분석하여 순환식 수경재배 시 스템 개발에 필요한 기초 자료를 확보하고자 본 연구를 수행하였다.
재료 및 방법
1. 실험장소 및 재료
충남농업기술원 과채연구소의 시설하우스 내에서 40일 간 육묘한 방울토마토(Lycopersicum esculentum Mill. cv. Minichal, Nongwoobio Co., Suwon, Korea) 묘를 정 식한 후 본 연구를 수행하였다.
2. 재배관리
2016년 3월 10일 펄라이트 배지가 충진된 재배 베드 에 재식거리 160×20cm로 조절하여 토마토 묘를 1주씩 정식하였고, 정식 직후부터 양액을 공급하였다. 본 연구 를 위한 양액은 야마자키 토마토 전용액을 사용하였으며 다량원소의 조성은 NO3-N 7, PO4-P 2, K 4, Ca 3, Mg 2meq·L-1, 미량원소는 Fe 2, B 0.2, Mn 0.14, Zn 0.02mg·L-1였다. 양액은 누적 일사량이 600W·m-2에 도달 할 때 1회 공급하였고, 관수 시간은 생육초기에는 2분으 로, 그리고 생육이 진전됨에 따라 총 공급된 양액의 약 30%가 배출되도록 공급량을 조절하였다. 배양액 EC의 경우 정식 후 활착기는 1.7dS·m-1로, 이후 생육 후기까지 2.0dS·m-1로 관리하였으며, pH는 5.5∼6.0을 목표로 질산 을 이용하여 조절하였다.
실험 기간 중 시설 내의 최저 온도가 13℃ 이하로 낮 아지면 온풍기가 자동으로 작동되도록 설정하였다. 측지 는 발생할 때마다 제거하였고, 10화방이 완전 개화한 후 상위 2엽을 남기고 적심하였다. 착과 유도를 위하여 화 방당 2~3개 꽃이 개화하였을 때 4-CPA(동부토마토톤, Farmhannong, Seoul, Korea)을 150배로 희석하여 분무 기로 살포하였다. 시험구는 완전임의배치 3반복으로 배 치하였으며, 재배 중 기타 관리는 충남농업기술원 과채 연구소 토마토의 재배방법에 준하였다.
3. 생육조사
초장, 절간장, 엽수, 엽면적, 엽장, 엽폭 등의 생육은 농촌진흥청 농업과학기술 연구조사 분석기준(RDA, 2003)에 따라 착과 절위별 화방의 2~3개 꽃이 50% 개 화된 시기에 반복당 10주씩 조사하였다. 또한 생육조사 시 식물체를 채취하여 잎과 줄기로 나눈 후 생체중을 측정하였고, 채취한 시료를 80°C 건조기에 48시간 건조 시킨 후 건물중을 측정하였다. 건물중 측정 후 분쇄기로 잎을 분쇄하여 무기성분 분석에 사용하였다.
4. 무기성분 분석
식물체 잎의 질소분석은 Kjeldahl(1030 analyzer, Kjeltec Auto, Jung-il Science, Hwaseong, Korea)법을 이용하였다. 시료 1g을 켈달플라스크에 평량하여 황산염 혼합분말과 진한 황산을 넣고 전기로에서 380℃, 80분간 분해하였다. 분해액은 증류수로 희석시킨 후 40% 수산 화나트륨을 가하여 산성을 알칼리화하여 증류한 다음 수 기는 2% 붕산을 사용하여 75~100mL 증류되면 0.05N 황산을 이용하여 종말점을 잡아 계산하였다. P, K, Ca 및 Mg의 분석은 시료 약 2g을 평량하여 삼각플라스크 에 취해, 질산 20mL과 과염소산 10mL를 넣고 혼합하 여 10시간 이상 정치 후 습식 분해하여 식힌 다음 다시 질산을 소량씩 가하면서 가열하여 미색이 되었을 때 분 해를 완료하였다. 분해가 끝난 시료는 질산을 이용하여 100mL로 정량하였고, 이 시료를 취하여 ICP(Integra XM2, GBC Scientific Equipment, Australia)를 이용하여 분석하였다(NIAST, 2000). 공급액과 배액의 무기원소 농도를 분석하기 위하여 생육조사 시기에 공급액과 배액 을 채취하였고, No. 2 여과지로 여과한 후 냉장 보관하 면서 분석에 이용하였다. 양액의 NO3-N는 Merck RQ flex(Eijkelkamp, Giesbeek, The Netherland)로, P, K, Ca, Mg 및 미량원소는 상기 분석방법과 동일한 ICP로 수행하였다.
결과 및 고찰
1. 방울토마토 착과 절위별 식물체 생육 및 무기성분 변화
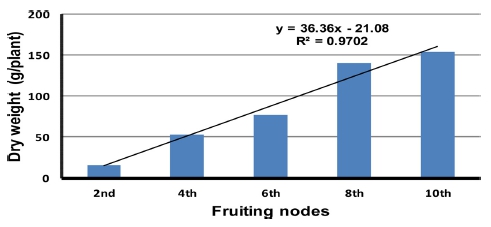
시험기간 중 각 착과 절위별로 화방의 꽃 중 2~3개가 약 50%정도 개화되었을 때 조사한 식물체 생육은 Table 1과 같다. 2번째 착과 절위의 화방이 개화하는 시기에 초장이 약 60cm였으며 착과 절위가 높아질수록 초장이 커져 10번째 착과 절위의 화방이 개화하는 시기에는 205cm까지 신장하였다. 10번째 화방이 개화하는 시기에 는 엽수 39.3매, 엽장 54.3cm, 엽폭 45.5cm로 2번째 화 방 개화시기와 비교하여 엽수는 2.5배, 엽장은 1.7배, 엽 폭은 2.1배 각각 증가하였다. 엽면적의 경우 2화방 개화 시기에 1,635cm2였지만 10화방 개화시기에는 6.6배가 증가된 10,272cm2 였다. Fig. 1은 각 착과 절위의 화방 이 개회시기 토마토 지상부의 무게 변화를 나타낸 것으 로 2번째 화방 개화시기에 식물체당 건물중이 약 16g였 으며 4화방 개화시기에는 3.3배 증가한 53g, 8화방 개화 시기에는 8.8배 증가한 140g, 10화방 개화시기에는 9.8 배 증가한 156g으로 무거워졌다.
Table 1. Growth characteristics of cherry tomato measured at flowering stage in each fruiting node.
| Fruiting node | Plant height (cm) | No. of leaves | Leaf length (cm) | Leaf width (cm) | Accumulated leaf area (cm2) |
|---|---|---|---|---|---|
| 2nd | 59.7 | 16.0 | 30.4 | 21 | 1,635 |
| 4th | 96.3 | 24.3 | 47.0 | 36.2 | 4,651 |
| 6th | 135.7 | 28.3 | 49.6 | 38.1 | 7,521 |
| 8th | 178 | 32.9 | 50.7 | 40.1 | 9,828 |
| 10th | 205.0 | 39.3 | 54.3 | 45.5 | 10,272 |
| LSDz | 9.9 | 1.0 | 2.2 | 4.3 | 574.8 |
| Linear | *** | *** | *** | *** | *** |
| Quadratic | *** | *** | *** | *** | *** |
각 착과 절위의 화방이 개화하는 시기에 식물체 잎을 수확하여 다량원소 함량을 분석한 결과는 Fig. 2와 같다. T-N 함량은 생육 초기인 2화방 개화기에 4.01%로 높았 으나, 5화방 개화기까지 점차 낮아졌고, 6화방 개화기에 는 3.21%로 5화방 개화기보다 소폭 증가하였다가 다시 10화방 개화기까지 감소하여 2.75%까지 낮아졌다. P함 량은 생육 초기인 2화방 개화기부터 5화방 개화기까지 점차 낮아졌다가 6화방 개화기부터 재배 종료기까지 점 차 증가하였고, 전 생육단계에서 0.28~0.35% 범위로 분 석되었다. K 함량은 T-N 함량과 반대의 경향을 보였다. 생육 초기인 2화방 개화기에는 2.01%로 함량이 적었으 나, 3화방부터 5화방 개화기까지 점차 낮아졌다가 이후 부터 높아져 10화방 개화기에는 2.83%였다. Ca 함량은 2화방 개화기에 2.07%로 가장 낮았지만, 생육 후기로 갈수록 함량이 높아졌으며, 특히 8화방 개화기에는 2.93%로 2화방 개화기보다 1.5배 높았다. Mg 함량은 전 생육기간 0.67~0.92%의 범위로 분석되었다.
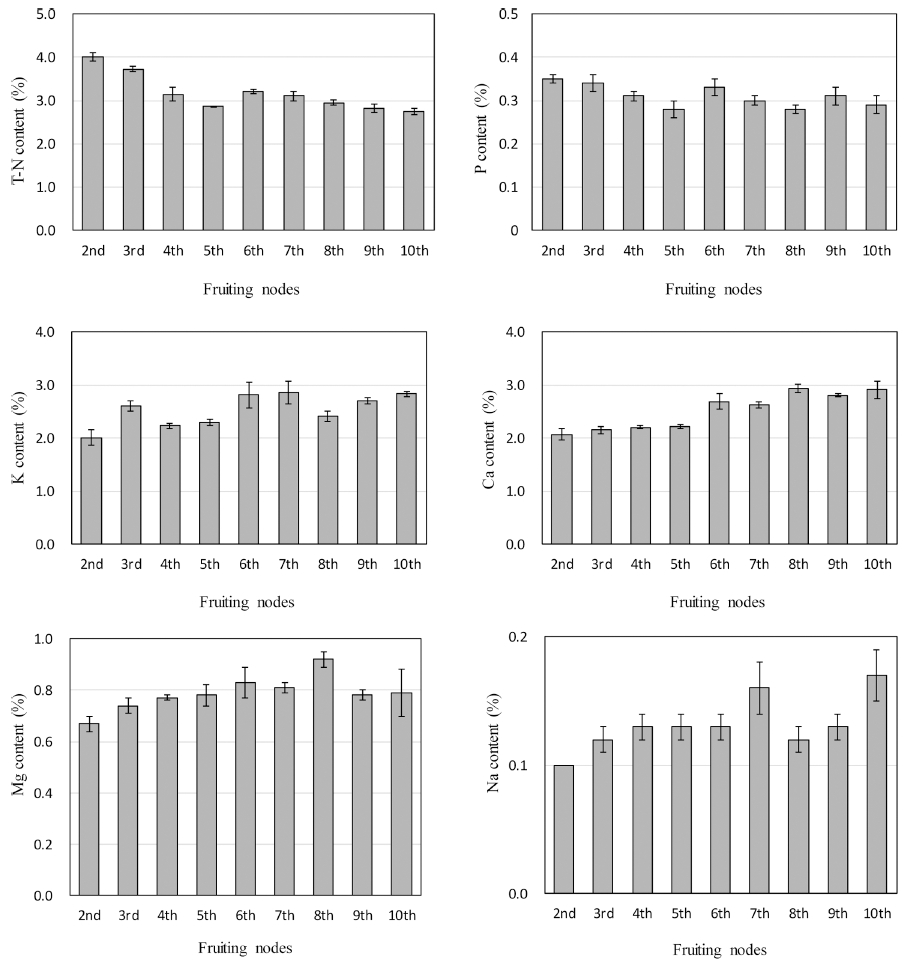
Sonneveld와 Voogt(2009)는 토마토와 오이를 재배하면서 신엽과 노엽의 무기원소 함량을 분석하여 보고하였다. 그 들은 노엽보다 신엽의 N 및 K 함량이 높고, Ca과 Mg 은 신엽보다 노엽의 함량이 월등히 높으며, P의 경우 노 엽보다 신엽의 함량이 높지만 큰 차이를 보이지 않았다 고 하였다. 이들은 각각의 작물 재배시기에 따라 노엽과 신엽을 분리하여 채취하였고, 본 연구에서는 각 착과 절 위에서 지상부의 잎 전체를 분석대상으로 삼은 차이가 있었다. 그러나 본 연구에서 착과 절위가 높아질 경우 총 수확한 잎이 많아질 뿐만 아니라 노엽의 비율이 증 가하였는데, 이러한 결과는 Sonneveld와 Voogt(2009)의 보고 내용 중 신엽에서보다 노엽과 더 유사한 무기물 함량을 보이며, 착과 절위가 낮고 식물체가 어린 상태에 서 채취한 잎은 신엽과 유사한 무기물 함량을 보이는 것으로 나타났다. 또한 Wilcox(1993)는 토마토를 정식한 후 105일까지 지상부 건물중 변화와 다량원소의 무기물 함량을 보고하였는데, 시간이 경과함에 따라 잎의 N, P, K, Ca 및 Mg 함량이 점차 감소하는 특징을 보였다고 하였다. Wilcox(1993)가 보고한 내용은 토경재배하면서 분석한 결과로써 고형배지경 수경재배를 한 본 연구와는 방법상의 차이가 있었다.
Sonneveld와 Voogt(2009)이 보고한 내용과 본 연구결 과를 비교할 때 질소를 제외한 P, K, Ca 및 Mg 함량이 상반된 결과를 보였고, Wilcox(1993)의 보고 내용과 비 교할 때는 N를 제외한 모든 무기원소에서 상반된 결과 를 보였다. 이와 같은 차이는 재배방법과 양액조성에서 원인을 찾을 수 있다. 본 연구를 위한 공급 양액은 NO3-N 7, PO4-P 2, K 4, Ca 3, Mg 2meq·L-1였고, 이를 mg·L-1의 농도로 환산하면 N 98, P 62, K 156, Ca 120 그리고 Mg은 48이다. 식물체 분석결과가 건물중에 기초 한 % 함량으로 나타내고 있으며, mg·L-1의 농도를 기준 으로 K 및 Ca이 N보다 더 많은 양이 시비되었고, 이러 한 이유로 양액의 EC에 더 큰 영향을 준 것으로 유추할 수 있다. 아울러 Fig. 3에 나타낸 배액의 전기전도도 (EC)와 Fig. 4에 나타낸 배액의 각종 무기원소 농도 변 화를 고려할 때 재배기간이 길어질수록 배지에 존재하는 K, Ca 및 Mg 등 양이온 농도가 대체적으로 높아졌는데 이는 결국 식물체의 흡수량 증가의 원인이 되었다고 판 단된다. 그러나 음이온인 NO3-N 및 PO4-P의 경우 매 관비시 쉽게 용탈되므로(Bar-Yosef, 2008) 배액의 농도 가 증가하였음에도 불구하고 배지 내 농도가 높아지지 않았고 결국 흡수량에 큰 영향을 미치지 않았다고 판단 하였다.
2. 공급액 및 배액의 무기성분 농도 변화
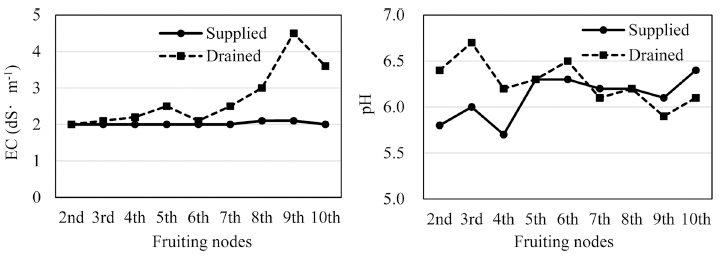
방울토마토 반촉성 수경재배 시 공급 양액 및 배출액 의 EC 및 pH 변화는 Fig. 3과 같다. 본 실험에서 활착 기 이후부터 생육 후기까지 양액의 EC를 2.0dS·m-1로 조절하여 공급하였음에도 정식부터 4화방 개화기까지는 배액의 EC가 큰 차이를 보이지 않았으며, 5화방 개화기 부터 높아지기 시작하여 9화방 개화기에는 배액의 EC가 4.5dS·m-1에 도달하였다. 5화방 개화기부터 EC가 높아진 것은 여러 연구된 보고와 유사하게(Seo et al., 2003; Zhang et al., 2010b) 재배시기가 여름철에 접어들면서 작 물의 수분 흡수량이 증가하기 때문인 것으로 생각된다.
Sonneveld와 Voogt(2009)에 의하면 물과 무기양분이 별도의 기작 하에 흡수되며, 각종 무기원소의 흡수 기작 또한 원소별로 다르다. 본 연구에서 동일하게 EC를 조 절하여 양액을 공급하였음에도 대기온도가 높아지는 시 기에 상대적으로 수분흡수량이 증가하였고, 이는 배지에 잔존하는 무기원소의 양이 증가하는 원인이 되어 배액의 EC가 점차 높아졌다고 판단된다. 아울러 배지에 양액을 공급하면 양액 속의 무기원소가 빠른 속도로 배지의 양 이온 치환 부위에 흡착되며, 양이온의 흡착량이 일정 수 준에 도달된 후에야 식물이 흡수할 수 있는 양으로 증 대된다(Bar-Yosef, 2008). 이 시기에 도달하면 배액의 무 기원소 농도도 높아지므로 본 연구의 재배 후기에 수거 한 배액의 EC가 높아진 또 다른 원인이 되었다고 생각 한다.
pH는 작물 생육에 있어 양분 흡수와 밀접한 관계가 있으며 대부분 원예작물의 경우 pH 5.5~6.8의 약산성을 좋아한다(Lunt and Clark, 1959; Yun et al., 2000). 본 실험에서 공급 양액의 pH를 5.6~6.4 범위로 조절하였음 에도 생육 초기에는 배액의 pH가 6.2~6.7로 공급액보다 높은 경향이었고, 7화방 개화기부터는 공급액보다 낮아 지는 경향이었다. 이는 Park과 Kim(1998)의 연구 보고 처럼 영양생장이 왕성할 때는 NO3-N 등 음이온의 흡수 량이 많고, 생식생장기에 K+ 등 양이온의 흡수가 증가하 여 뿌리로부터 수소이온(H+)을 방출하기 때문인 것으로 생각된다.
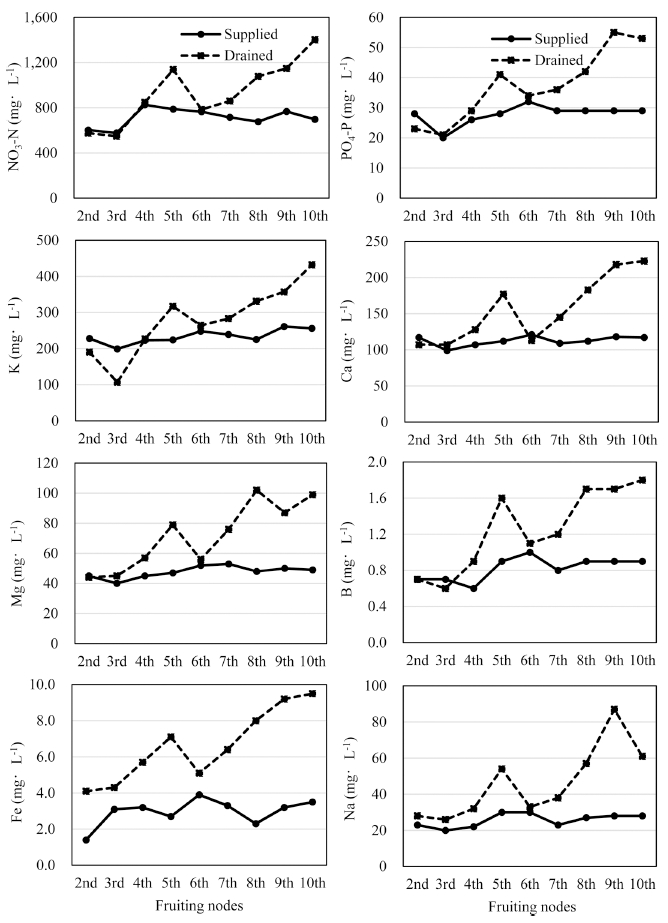
공급액 및 배액을 채취하여 NO3-N, P, K, Ca, Mg 등 을 분석한 결과는 Fig. 4와 같다. NO3-N과 P는 생육초 기인 4화방 개화기까지 공급액 및 배액의 농도가 유사 하였으나, 5화방 개화기 이후에는 공급액보다 배액에서 높은 농도로 분석되었고, 특히 9화방 개화기에는 공급액 보다 2.0~2.3배 높았다. K는 3화방 개화기에는 배액의 농도가 낮았으나 5화방 개화기 이후에는 NO3-N 및 P와 유사하게 높아지는 경향이었다. Ca 및 Mg는 2, 3 및 6 화방 개화기에는 공급액과 배액의 농도 차이가 크지 않 았으나 각각 5화방 및 7화방 개화기 이후에는 높아지는 경향을 보였다. B와 Fe 역시 착과 절위별로 공급액보다 배액에서 농도가 높았으며, 특히 5, 8, 9, 10화방 착과 절위에서 채취한 시료액은 1.5~3.5배 가량 높았다.
전반적으로 본 실험의 연구 결과로 미루어 보아 공급 액보다 배액에서 무기성분 농도가 높은 경향을 나타내었 고, 특히 5화방 및 9화방 착과 절위에서 채취한 시료의 농도가 높았다. 이는 고온기로 변화되면서 작물체의 수 분 흡수량이 증가하여 배출되는 물량이 적어지고 배지 내에 집적된 무기원소의 양이 증가한 주요 원인과 함께 앞에서 설명한 요인들이 작용하였기 때문인 것으로 판단 된다. Zhang et al.(2010a)도 파프리카의 생육시기별로 배액의 무기원소 농도를 분석하였을 때, 생육 후기에 무 기성분 농도가 급격히 높아졌다고 하였다. Kim et al.(2001)도 작물이 고온기에 수분을 많이 흡수하기 때문 에 배지의 무기이온 농도가 높아지며 무기이온 농도 변 화는 일사량과 상관관계가 높다고 보고한 바 있다.
